

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Gastric cancer is one of the leading fatal malignancies worldwide (1), and the most common malignancy in Korea (2). Now, gastric cancer has been reported to be the second leading cause of death due to malignancy in Korea, which was the first leading cause before 2005 (2). However, the etiology of gastric cancer has not yet been unveiled completely, and the previous studies have provided evidence that risk factors such as diet, infection including Helicobacter pylori and Epstein-Barr virus, and hereditary trait may be involved in (3). Recently, accumulation of molecular events has been known to be important in the development of gastric cancer (4). Furthermore, these biological markers as well as traditional histopathological parameters can be used as prognostic factors (4-8).
It was suggested that beta-catenin can regulate NF-kappaB negatively (9), and our previous study revealed that NF-kappaB nuclear positivity was more frequent in Epstein-Barr virus (EBV)-positive gastric carcinomas than conventional gastric carcinomas (10).
Beta-catenin, a 92-kDa protein (11), plays multifunctional roles. First, beta-catenin is a component of the cell-cell adhesion complex, as an E-cadherin/beta-catenin complex. Secondly, beta-catenin acts as a coactivator of transcription factors involved in the Wnt signaling pathway (11). Wnt signaling is mediated by secreted proteins that interact with specific cell-membrane receptors and is involved in the growth and differentiation of cells during development. Wnt pathway dysregulation has been implicated in many cancers (11). Uncontrolled activation of this signaling pathway may induce inappropriate proliferation of target cells and may contribute to the development of malignancy. Dysfunction of this regulatory pathway may result in the accumulation of a hypophosphorylated stable form of beta-catenin in the cytoplasm (11), from whence it translocates to the nucleus, binding to the high mobility group domain factors Tcf/LEF (12) and stimulating transcription of target genes such as c-myc and cyclin D1 (13).
E-cadherin is a 120-kDa transmembrane glycoprotein that is responsible for calcium-dependent intercellular adhesion by homotypic interactions (14), and hence plays a critical role in cancer cell metastasis and invasion because anchorage of cells to substrate is critical for the integrity of many cell types including epithelial cells. Becker et al. first disclosed that E-cadherin is implicated in gastric carcinogenesis (15). Functional cadherin-dependent cell adhesion requires the formation of complexes between E-cadherin and catenins. However, cell growth suppressor activity of E-cadherin is not adhesion-dependent. E-cadherin protein can directly transduce a growth inhibitory signal, through the inhibition of beta-catenin signaling (16).
Previous immunohistochemical studies have shown a close relationship between alteration of E-cadherin or bata-catenin and lower 5-yr survival rates in diverse human cancers such as breast, urinary bladder, prostate, and esophagus cancer (17-20). However, there is a conflict concerning the prognostic significance of bata-catenin or E-cadherin in gastric cancer (5-8, 21-23).
In the present study, we investigated in situ hybridization for EBV-encoded small RNAs and immunoexpression of beta-catenin and E-cadherin proteins in 111 cases of gastric carcinomas, in order to define a relationship between beta-catenin and EBV and to clarify the impact of beta-catenin or E-cadherin on the prognosis.
MATERIALS AND METHODS
Patients and tumor tissue
The present study was based on data from 111 Korean patients undergoing surgically resection for gastric carcinomas from 1990 to 1994 at the Seoul National University Boramae Hospital. Age, sex, tumor size and location, tumor differentiation according to the WHO classification, Lauren classification, and pathological tumor stage (TNM stage) according to the American Joint Committee on Cancer (AJCC) system were evaluated. Survival data were obtained from hospital records and patients' physicians in charge. The overall survival period of the patients was measured from the date of operation to the date of death or the last follow-up.
EBER-in situ hybridization
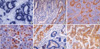
Three micromete thick sections were cut from each paraffin block, deparaffinized and dehydrated, digested with proteinase K, and hybridized for two hours at 37℃ with a fluorescein-conjugated EBV oligonucleotide probe targeting EBV-encoded RNA (EBER) (Novocastra, Newcastle-upon-Tyne, U.K.). Hybridization products were detected using an alkaline phosphatase-conjugated antibody to fluorescein isothiocyanate (affinity-isolated rabbit F[ab']). 5-Bromo-4-chloro-3-indolylphosphate and nitroblue tetrazolium were used as an enzyme substrate to demonstrate the alkaline phosphatase activity. The slides were counterstained with Mayer's hematoxylin. Using a light microscope, black or dark navy-colored aggregates at the hybridization site were interpreted as positive staining (Fig. 1A). Rare reactive lymphocytes showed a positive signal of EBER, but only signals of EBER within the tumor cells were considered to be positive.
Immunohistochemistry
Tumor tissue was retrieved from formalin-fixed, paraffin-embedded specimen. After the antigen retrieval process by microwave, immunohistochemistry was performed with monoclonal antibodies against E-cadherin (1:100, Zymed Laboratories, San Francisco, CA, U.S.A.) and beta-catenin (1:100, Transduction Laboratories, Lexington, KY, U.S.A.), using avidin-biotin complex method. For an interpretation of immunohistochemical staining, cut-off point was adapted as reported previously (7, 21, 22, 24-26). Those cases with loss of membranous staining in more than 10% of tumor cells were considered to be an altered expression of E-cadherin protein (7, 24, 25). An altered expression of beta-catenin protein was divided into three patterns, that is, loss of membranous staining in more than 10% of tumor cells (7, 24, 25), cytoplasmic expression in any degree of tumor cells (21, 22, 26) and nuclear accumulation in any degree of tumor cells (21, 22, 26) patterns.
Statistics
All statistical analyses were conducted using the SPSS 10.0 statistical software program (SPSS, Chicago, IL, U.S.A.). The chi-square test or Fisher's exact test (two-sided), Pearson correlation (two-tailed), and Kendall's tau-b correlation were applied. The cumulative survival rates were obtained by the Kaplan-Meier analysis, and differences in survival were compared using the log-rank test. Multivariate analysis was performed using the Cox proportional hazards model. Values of p<0.05 were considered to be statistically significant.
RESULTS
Cilnicopathological features
Clinicopathological features are summarized in Table 1 and 2. The follow-up period ranged from 2 to 167 months, with a median of 50 months; thirty-five patients (31.5%) were dead and 76 (68.5%) were alive. Among the clinicopathological features evaluated in the present study, Lauren classification showed correlations with tumor location, lymph node metastasis (pN stage), TNM stage, and patient's survival (p<0.05).
EBV-infected gastric carcinomas
EBV infection was revealed in seven carcinomas (6.3%) (Fig. 1A), and none of EBV-infected gastric carcinomas showed beta-catenin nuclear accumulation. However, five out of seven EBV-infected carcinomas revealed beta-catenin alteration; three cases showed cytoplasmic expression, and two out of these three cases had loss of membranous staining simultaneously. Another two cases revealed loss of membranous staining. Regarding E-cadherin, only one EBV-infected carcinoma disclosed E-cadherin alteration. However, there were no statistical significances in beta-catenin or E-cadherin immunoexpression patterns between EBV-infected gastric carcinomas and non-infected carcinomas (Table 3).
The EBV infection status showed a marginal impact on the patient outcome (p=0.08) (Table 2) but was not associated with beta-catenin or E-cadherin alteration.
Expression of beta-catenin and E-cadherin
There were no significant relationships between clinicopathological features and immunoexpresssion of beta-catenin or E-cadherin (Table 2, 3). A representative immunohistochemical staining pattern is shown in Fig. 1B-F. In normal gastric epithelium, both E-cadherin and beta-catenin showed cytoplasmic membranous staining (Fig. 1B, E). Eighty cases (72.1%) showed beta-catenin alteration, i.e. loss of membrane staining in 65 cases (58.6%), cytoplasmic expression in 35 cases (31.5%), and nuclear accumulation in 15 cases (13.5%) (Table 1, 2). Regarding concurrent cytoplasmic and nuclear beta-catenin expression, nine of 35 cases displayed both cytoplasmic and nuclear beta-catenin. In six additional cases (5.4%), beta-catenin was seen only in the nuclei, leading to a total of 15 cases (13.5%) with beta-catenin nuclear accumulation. E-cadherin alteration (loss of membrane staining) was observed in 34 cases (30.6%) of gastric carcinomas.
The E-cadherin alteration was statistically significantly correlated with beta-catenin alteration (p=0.003); in particular, with beta-catenin membranous loss or nuclear accumulation (p=0.012 and p=0.040, respectively) (Table 4).
Survival analyses
The beta-catenin nuclear accumulation was associated with a poor patient survival in a subset of Lauren diffuse-type carcinomas. Additionally, in a subset of male patients, beta-catenin nuclear accumulation and E-cadherin alteration were related to a poor patient survival (p<0.05).
The combined expression pattern of beta-catenin/E-cadherin was reclassified into four combinations (considering Table 4); the combination of beta-catenin alteration/E-cadherin, the combination of beta-catenin membranous pattern/E-cadherin, the combination of beta-catenin cytoplasmic pattern/E-cadherin, and the combination of beta-catenin nuclear pattern/E-cadherin. On univariate analysis, the combination of beta-catenin nuclear pattern/E-cadherin had a significant impact on the patient survival (p<0.05) (Fig. 2A). In other words, a group with beta-catenin nuclear accumulation/E-cadherin alteration showed poor patient survival. Elaborating on this, in groups of combined beta-catenin nuclear pattern/E-cadherin, the patient survival rate of each group was 63% in beta-catenin nuclear accumulation/E-cadherin alteration, which contrasted with the remaining groups; 29%, 29%, and 21%, in a group with beta-catenin nuclear accumulation/normal E-cadherin, in a group with beta-catenin, no nuclear accumulation/normal E-cadherin, and in a group with beta-catenin, no nuclear accumulation/E-cadherin alteration, respectively (Fig. 2A).
On multivariate analysis, the combined beta-catenin nuclear pattern/E-cadherin, and cancer stage (TNM classification based on the AJCC system) had significant impacts on patient survival (p=0.008 and p=0.001, respectively) (Table 5, Fig. 2). Besides, beta-catenin nuclear accumulation showed a marginal impact (p=0.077) (Fig. 2C).
DISCUSSION
In the present study, we suggest that beta-catenin activation through nuclear accumulation seldom occurs in EBV-infected gastric carcinomas, although beta-catenin expression can be altered in patterns of membranous loss and cytoplasmic expression. Furthermore, our previous paper showed that NFkappaB nuclear accumulation occurred more frequently in EBV-infected gastric carcinomas than in EBV-negative gastric carcinomas (p<0.05) (10), suggesting EBV-infection could be related to NFkappaB activation. The beta-catenin cytoplasmic stabilization is involved in the first step of beta-catenin activation, and nuclear transport is the second step (27), In the present study, it was not certain whether cytoplasmic expression might be related to activated form of beta-catenin or not, because we have not evaluated the free beta-catenin level in cytoplasm or beta-catenin-dependent gene expression level. However, beta-catenin nuclear accumulation can represent beta-catenin activation. Consequently, in EBV-infected gastric carcinomas, beta-catenin activation through nuclear accumulation hardly occurs, but NF- kappaB could be activated. It is plausible that NFkappaB activation might suppress beta-catenin activation in EBV-infected gastric carcinomas because previous papers suggest that beta-catenin activation can inhibit NFkappaB activation (9). Therefore, a negative crosstalk between beta-catenin and NFkappaB activation seems to exist in EBV-infected gastric carcinomas.
In the present study, E-cadherin alteration (loss of membrane staining) was observed in 30.6% and beta-catenin alteration in 72.1% [loss of membrane staining in 58.6%, cytoplasmic expression in 31.5%, and nuclear staining in 13.5%] of gastric carcinomas. The figures of the E-cadherin alteration and beta-catenin nuclear accumulation are almost same as those in the paper by Grabsch et al., 30.7% and 13.2%, respectively (21). Additionally, the figures of the beta-catenin membranous loss, cytoplasmic expression and nuclear accumulation are similar to those in the paper by Nabais et al. (22, 26). Meanwhile, concerning beta-catenin, some studies did not evaluate nuclear accumulation of beta-catenin (5, 6, 8, 23, 28). However, beta-catenin nuclear accumulatin should be evaluated in the study of the Wnt signal activation because beta-catenin translocated into the nucleus is predominantly in an active and oncogenic form (11, 27, 29, 30). The Wnt signal activation induces beta-catenin stabilization and its accumulation in the cytoplasm, and then, beta-catenin is translocated to the nucleus, where it interacts with DNA binding proteins of the T-cell factor-lymphoid enhancer factor (Tcf/LEF) family, causing transcriptional activation of target genes (11, 27, 29, 30), such as c-myc and cyclin D1 oncogenes (13). Moreover, in the present study and others (26), certain cases showed beta-catenin only in the nuclear accumulation pattern, neither in cytoplasmic expression nor membranous loss. Therefore, beta-catenin nuclear accumulation should be assessed necessarily.
In the multivariate survival analysis, a group with combined beta-catenin nuclear accumulation/E-cadherin alteration showed a poor patient survival, compared to the other groups (Fig. 2A). In this combined pattern, oncogenic process by beta-catenin activation and cancer progression due to reduction of the cell-cell adhesion by E-cadherin loss can occur together and end up with a poor patient survival, accordingly. This result might apparently support previous suggestions that the beta-catenin or E-cadherin immunoexpression pattern correlates with patient survival (5-8), but the present study showed that only beta-catenin nuclear accumulation or only E-cadherin alteration had no significant impact on patient survival, or rather a combined pattern of beta-catenin nuclear accunmulation/E-cadherin alteration was an independent prognostic factor.
There have been controversies concerning the prognostic significance of E-cadherin or bata-catenin in gastric cancer; that is, Grabash et al. (21), Nabias et al. (22), and Joo et al. (23) showed no prognostic significance in their studies. This discrepancy might be partly due to the interpretation of immunohistochemical staining or case selection bias. First, we can find a fault with ignorance of beta-catenin nuclear accumulation in some papers (5, 6, 8, 23, 28). Secondly, subsets of cases can make wide differences. For example, in the present study, beta-catenin nuclear accumulation or E-cadherin alteration had a significant impact on patient survival in subsets such as Lauren diffuse-type patients and male patients, but a marginal impact in the total patient group as a whole. Additionally, Nabais et al. demonstrated that beta-catenin or E-cadherin did not have a prognostic significance based upon different subsets of gastric carcinomas (22).
Previous studies analyzed the prognostic significance of each protein, beta-catenin or E-cadherin, rather than the combined pattern of beta-catenin/E-cadherin. To the best of our knowledge, there is a study about the combined expression pattern of beta-catenin/E-cadherin, in which it correlated with the patient survival in univariate analysis, but not in multivariate analysis (23), although it could be a critical drawback that beta-catenin nuclear accumulation was not analyzed in their study (23). Besides, the other combined pattern of abnormal E-cadherin/MUC1-positive was suggested to be an unfavorable prognostic factor (8).
In conclusion, beta-catenin expression can be altered in EBV-infected gastric carcinomas, but beta-catenin activation through nuclear accumulation hardly occurrs. The beta-catenin and E-cadherin alterations correlated with each other. The combined beta-catenin and E-cadherin pattern can be used as prognostic markers in Korean gastric carcinomas, and frequent alterations of beta-catenin and E-cadherin proteins are observed.






 XML Download
XML Download