

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Chimeras have been defined as individuals whose cells are derived from two or more zygotes (1). Several different types of chimeras exist (2, 3): artificial chimerism, twin chimerism, and dispermic chimerism. These forms of chimerism are restricted to hematopoietic tissue while dispermic chimeras demonstrate a dual-cell population in disparate body tissues derived from different germ layers (3, 4).
Chimerism in humans is a rare phenomenon usually identified at the time of blood group determination when mixed field agglutination on the cell typing is noted (3). A report of a hemolytic transfusion reaction related to the transfusion of chimeric red blood cells (RBCs) has recently been published (5).
A few cases of phenotypically normal dispermic chimeras have also been reported (1). The 47,XYY syndrome is characterized by a male karyotype with an extra copy of the Y chromosome. In most cases this chromosomal abnormality causes no unusual physical features or medical problems, especially in men with a mosaic 46,XY/47,XYY karyotype (6), hence many males with this syndrome are undetected.
We report a case of dispermic chimerism in a 39-yr old Korean blood donor who presented with mixed field agglutination in his B antigen typing, and a mosaic 46,XY/47,XYY karyotype.
CASE REPORT
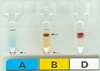
The propositus was a 39-yr old father of one child, healthy, morphologically normal Korean male who demonstrated mixed field hemagglutination on cell typing by both the manual tube and gel card techniques at the time of blood donation (Fig. 1). This mixed field agglutination pattern is consistent with an ABO subtype that is relatively common in Korea known as B3 and is caused by a series of mutant B alleles known collectively as B3 alleles.
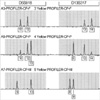
To investigate the hypothesis that the mixed field agglutination in the propositus was caused by a B3 allele, allele-specific polymerase chain reaction (AS-PCR) and direct sequencing of exons 6 and 7 of the ABO gene were performed using previously described methods (7, 8) on the propositus' genomic DNA. Initially these investigations demonstrated only the O01/O02 genotype. However, a careful review of these chromatograms revealed very small B-allele specific peaks at nucleotides 526, 657, 703, 796, 803, and 930 (Fig. 2). The guanosine residue at position 261 was not detected. The former single nucleotide polymorphisms (SNP) discriminate common O and A alleles from B alleles, while the 261delG SNP is only present in common O alleles.
In an effort to demonstrate a B allele in the propositus, haplotype-specific templates were prepared using B allele-specific primer pairs. Haplotype sequencing revealed the presence of a normal B101 allele. This normal B101 allele was also detected when exons 6 and 7 of the propositus' ABO gene were cloned and sequenced. Thus 3 ABO alleles, B101, O01 and O02, were conclusively demonstrated to be present in the propositus. This confirmed that the mixed field agglutination observed with anti-B reagent indicated the simultaneous presence of two distinct RBC populations (group B and group O) in the propositus.
The propositus denied receiving a stem cell transplant or transfusion and did not have a known twin, thus chimerism or mosaicism was suspected to explain the presence of the 3 ABO alleles.
To discriminate chimerism from mosaicism, nine short tandem repeat (STR) loci were tested on DNA extracted from blood, buccal mucosal cells, and hair from the propositus, and on DNA isolated from blood on his immediate relatives (father, mother, and older brother). Out of 9 STR loci, 4 loci (D3S1358, D5S818, D13S317, and D18S51) demonstrated a pattern consistent with a double paternal DNA contribution confirming the presence of dispermic chimerism (Fig. 3). To investigate the extent of the propositus' chimerism, ABO typing as well as sequence analysis of ABO exon 6 and 7 was performed on his immediate relatives (Table 1). Both parents were blood type B and shared the B101 allele while the O01 allele was also detected in the propositus' mother, and the O02 allele in his father. His brother was also had blood type B with the B101/O02 genotype.
Using sequence-based typing on genomic DNA, one paternal and one maternal HLA class I (A, B) and II (DRB1) haplotype was identified in the propositus (Table 1). A double paternal DNA contribution was thus not identified. Analysis of 50 metaphases indicated that the propositus was also a karyotypic mosaic: 32 (64%) metaphases exhibited a normal male karyotype, 46,XY, while 18 (36%) metaphases demonstrated 47,XYY. No structural chromosomal abnormalities were present.
DISCUSSION
One of the common presentations of morphologically normal appearing chimeras is an unusual finding in the ABO typing such as mixed field RBC agglutination. However, mixed field agglutination can be seen in numerous situations including ABO-incompatible stem cell transplantation, rare ABO subgroups such as B3, recent transfusion of type-O RBCs in a non-O recipient, twin chimerism, dispermic chimerism, and mosaicism. Most of these entities can be identified by investigating the individual's clinical and transfusion history or by conducting additional routine serologic tests and/or molecular tests including ABO genotyping (9).
Because this healthy and fertile propositus neither had a history of therapeutic transfusion/stem cell transplantation nor knowledge of a twin, we initially suspected a fairly common (amongst Koreans) group B subtype that classically presents with mixed field agglutination, B3, as the cause of his unusual cell typing. The unexpected absence of a conclusive B allele by routine ABO sequencing led us to suspect the presence of chimerism, and more sensitive ABO sequencing techniques were required to reveal a B allele. In the end, a normal B101 allele, in addition to two common O alleles, were discovered in the propositus. Assuming that the B101 allele was inherited from his father, the propositus' ABO genotype is most likely B101/O01 and O01/O02. Thus his RBC mixed field agglutination pattern was not due to a B subtype allele, but to the simultaneous presence of group B and O RBCs. As blood group genotyping becomes the mainstream and genotype/phenotype discrepancies are detected more commonly, more cases of chimerism might be detected and thus its true incidence might be uncovered.
To determine if the propositus was a chimera or a mosaic, DNA polymorphisms from tissues derived from different germ layers were analyzed. DNA from the propositus' hair and buccal mucosa (ectodermally derived) and his blood cells (mesodermally derived) were tested, and at four loci revealed a pattern consistent with a double paternal DNA contribution (Fig. 3). The finding of a double paternal DNA contribution in these disparate tissues indicates that the propositus is most likely a dispermic chimera resulting from parthenogenetic division of the ovum and its subsequent fertilization by two spermatozoa carrying different paternal DNA-one with a 23,Y chromosome complement and another spermatozoa with a different 24,YY DNA complement (10). This would explain the apparent single maternal DNA contribution. Alternatively, a non-disjunction event producing the mosaic 47,XYY karyotype could have occurred after fertilization with two different 23,Y spermatozoas. HLA typing failed to reveal the true number of parental loci present. This was probably due to a number of factors including the relatively low copy number in the peripheral blood of the second paternal DNA contribution, and to the relative insensitivity of whole blood PCR-based techniques for detecting small quantities of DNA. In other words, the same reasons we initially failed to detect the B allele from peripheral blood DNA were probably the same reasons we did not detect a double paternal HLA contribution. A similar phenomenon was also reported by Drexler et al. (11) in their tetragametic chimera, only a single paternal HLA haplotype was identified, while both maternal haplotypes were present.
The propositus' karyotype revealed a mosaic pattern with 32/50 metaphases demonstrating 46,XY and 18/50 demonstrating 47,XYY. 47,XYY is a common chromosomal disorder found in approximately 1/1,000 males (6). Of all cases of 47,XYY syndrome, 80-90% are 47,XYY and 10-20% are 47,XYY/46,XY (6). As with our propositus, the majority of 47,XYY males are phenotypically normal and fertile, bearing chromosomally normal offsprings (12). This case represents the first description of a morphologically normal and fertile dispermic chimera with mosaic 47,XYY syndrome.


 XML Download
XML Download