

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Centric fission results when a metacentric or submetacentric chromosome splits at the centromere, giving rise to two stable telocentric products, isochromosomes, or ring chromosomes. Stable fission chromosomes in human were first described by Sinha et al. (1) and have subsequently been observed in several pedigrees (2, 3), in a few individuals (4-8), and in amniotic fluid cultures (7, 9). The heterozygous individuals may have a balanced complement of genetic material and thus be phenotypically normal. But they appear to be at increased risk for production of unbalanced gametes which consequently increased risks for spontaneous abortion, still-birth and live born infants with clinical effects from unbalanced genotypes.
In this paper we report the first case of de novo centric fission of chromosome 11 detected in blood lymphocyte chromosome analysis.
CASE REPORT
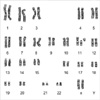
A 25-yr-old female patient visited the habitual abortion clinic in Hanyang University Hospital because of her history of recurrent abortions. She had been pregnant 3 times and all pregnancies were resulted in spontaneous abortions during 8-10 weeks of gestation. All of the 30 metaphases analyzed showed karyotype of 47,XX,-11,+fis(11)(p10),+fis(11)(q10) (Fig. 1). Both arms of the broken chromosome 11 appeared to be telocentric chromosomes. Chromosome studies on all her family members (father, mother and a younger brother) revealed normal karyotypes.
Six months later, she became pregnant. A prenatal cytogenetic test was performed by chorionic villus sampling at 10th week of pregnancy. The fetus had normal female karyotype, 46,XX, and she delivered a healthy female baby. The karyotype of the baby was reconfirmed by peripheral blood chromosome analysis.
Cytogenetic analysis
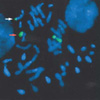
Chromosome analysis of 72 hr lymphocyte cultures was performed with conventional GTG-banding techniques. To confirm the chromosome aberration of fis (11), primed in situ labeling (PRINS) with the oligonucleotide primer specific for centromere of chromosome 11 (9) and fluorescence in situ hybridization (FISH) with whole chromosome painting probe for chromosome 11 (Chromoprobe TM-M11, Cytocell, Cambridge, U.K.) were studied. In all the metaphases scored (more than 30), 47 chromosomes were found. Chromosome 11 was divided into one smaller and one larger telocentric chromosome (Fig. 1). FISH analyses showed that no other chromosomes were involved (Fig. 2). It was evident that both fragments possessed centromeric heterochromatins in different amounts of the FISH positive band, i.e. the telocentric 11p has much larger centromeric heterochromatin than that of the telocentric 11q (Fig. 3). Centric fission of chromosome 11 has resulted in two stable telocentrics. There was no tendency to fuse again or to associate with acrocentrics as verified in 50 additional metaphases. Three distinct fluorescent spots were detected with the PRINS reaction in more than 100 interphase nuclei.
DISCUSSION
Individuals with stable centric fission products have rarely been documented in human, and to our knowledge this is the first report on centric fission of chromosome 11. In this case, the centric fission does not cause any significant dysmorphic features and mental handicap except for reproduction failures.
For many years, karyotype evolution was thought to occur via chromosome fission, fusion, translocation, and inversion. Both centric fission and centric fusion have been considered as an explanation of the diverse range of chromosome numbers among species (11).
Two major mechanisms have been proposed for the generation of chromosome fission (12, 13). One is the simple centric fission which is a simple transverse split at the centromere to result in two stable C-banded and α-satellite-positive centric fission products. The other mechanism is more complex including the pre-duplication of a centromere or the activation of a new centromere. To retain a mitotically stable status of fission products, the end may be protected by adding telomeric sequences or recombination resulting in a formation of isochromosome or ring chromosome (14, 15).
In our case, the fission products were apparently generated by simple centric fission mechanism. In PRINS result, the signals of the fission products were smaller than the signal of the normal chromosome 11. This may support the simple centric fission mechanism in our case. However, even between homologous chromosomes, the size of centromeres is different from each other, so a further study is required to understand the detail mechanism of this case.

 XML Download
XML Download