

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Orientia tsutsugamushi causes scrub typhus, which is endemic in many countries in the Asia-Pacific region, including Korea (1, 2). This disease is an acute febrile illness characterized by fever, rash, and eschar. The mechanism of its pathogenesis has not been studied extensively, but is believed to be similar to that of other Rickettsia, which includes endothelial damage and subsequent vasculitis (3).
Doxycycline has been used as the drug of choice for scrub typhus. However, recent reports of doxycycline-resistant strains in Thailand have stimulated the research on alternative antibiotics for the treatment of this disease (4). Because the incidence of scrub typhus has recently increased markedly, there is growing concern about the possible occurrence of a resistant strain in Korea. Therefore, it is necessary to test the susceptibility of Korean isolates of O. tsutsugamushi to doxycycline and other antibiotics (5).
The testing of O. tsutsugamushi antibiotic susceptibility has not been standardized because it requires viable host cells for bacterial growth. In a previous study, we reported the characteristics of a monoclonal antibody (MAb), M686-13, that reacts exclusively with intracellular O. tsutsugamushi, and its application in an immunofluorescence assay (IFA) to determine antibiotic susceptibility (6, 7). However, that method requires the manual count of bacterial particles or computer-assisted scanning. Therefore, we were interested in finding an objective and convenient method with which to accurately quantify O. tsutsugamushi.
Flow cytometry has been used previously for antibiotic-susceptibility testing of intracellular pathogens (8-13). A pioneering work by Kelly et al. described the use of flow cytometry to measure the growth of O. tsutsugamushi in cells treated with antibiotics (14). Although their method is relatively rapid and objective, it is not sufficiently sensitive to distinguish quantitatively between low levels of infection. Moreover, it may be less sensitive for the detection of diverse local strains that are antigenically heterogeneous (14). We speculated that low sensitivity resulted from the use of polyclonal animal serum to stain the bacteria. Therefore, we decided to improve the efficiency of flow cytometric detection by using a specific MAb.
In this study, we demonstrate an improved flow-cytometric approach to the sensitive and quantitative measurement of the growth of O. tsutsugamushi. We used our method to determine the doxycycline susceptibility of strains from Korea and Thailand.
MATERIALS AND METHODS
Cell culture and O. tsutsugamushi
Cells of the human umbilical-vein-derived endothelial cell line ECV304 were maintained in M199 medium (Gibco BRL, Gaithersburg, MD, U.S.A.) supplemented with 10% (v/v) heated-inactivated fetal bovine serum (Gibco BRL) in a humidified atmosphere containing 5% CO2 at 37℃. The Boryong serotype of O. tsutsugamushi (15) was cultivated in ECV304 cells, as described previously (16). The Thailand strains (AFSC-4, AFSC-7, TA686, TA678, TA716, TA763, and TH1817) were kindly provided by Dr. Daniel Strickman, Naval Medical Research Institute, U.S.A.. The Karp (ATCC VR-150) and Gilliam (ATCC-VR-312) strains were obtained from American Type Culture Collection; Kato strain was donated from Dr. Hiroshi Tanaka, the Institute of Medical Science, Tokyo University, Japan. Kuroki and Kawasaki strains were donated from Dr. Akira Tamura, Department of Microbiology, Niigata College of Pharmacy, Japan. All bacterial strains were cultivated in ECV304 cells. When infected ECV304 cells showed maximum cytopathic effects, the cells were disrupted with glass bead (diameter 1.0 mm) and centrifuged at 300×g for 5 min. The resulting supernatants were immediately used to infect further ECV304 cells.
Treatment with doxycycline
ECV304 cells grown in 6-well plates were incubated with O. tsutsugamushi for 3 hr, allowing time for O. tsutsugamushi to attach to and enter the host cells. At the end of the initial incubation period, the inoculums were replaced with fresh medium containing two-fold dilutions of doxycycline (Sigma, St. Louis, MO, U.S.A.) from 0.2 µg/mL to 0.00625 µg/mL, and the cells were then re-incubated in 5% CO2 at 37℃ for three days. Doxycycline was prepared in aliquots of 0.5 mL at an active concentration of 5,000 µg/mL in sterile distilled water. These aliquots were stored frozen at -20℃ until required.
Monoclonal antibodies and selection of antibodies for the application in flow cytometry
MAb FS15 and FS10 react against a linear epitope on 56-kDa major outer membrane protein of O. tsutsugamushi (17). Other MAbs (Rb167, Rb134, M686-8, and Shim107) were obtained from cell fusion experiments of spleen cells of mice immunized with live O. tsutsugamushi, as described previously (6, 18). For its application to flow cytometry, MAb FS15 was purified on an AffinityPak Protein A Column (Pierce Biotechnology Rockford, IL, U.S.A.) and conjugated with fluorescein isothiocyanate (FITC) with EZ-label Fluorescein Protein Labeling Kit (Pierce Biotechnology), according to the manufacturer's instruction.
To select a suitable MAb for the application in flow cytometry, we tested the immunoreactivities of MAbs to various strains of O. tsutsugamushi using IFA. Among the tested MAbs, we selected the MAb that had broad reactivity to many strains and stained the bacteria brightly.
Immunofluorescent antibody staining
The reactivities of MAbs to various strains of O. tsutsugamushi were examined using IFA. ECV304 cells infected with each strain were fixed with acetone-methanol (1:1) or acetone and treated with dilutions of MAbs in phosphate-buffered saline (PBS) for 1 hr at 37℃. After the cells had been washed briefly with PBS, they were treated with FITC-conjugated goat anti-mouse IgG (Jackson ImmunoResearch, West Grove, PA, U.S.A.) for 1 hr in a moist chamber, and then washed three times with PBS. To clearly define the O. tsutsugamushi, the host cells were counter-stained with 0.003% Evans blue in PBS. After the slides had been washed, they were placed on mounting medium (Vector Laboratories, Burlingame, CA, U.S.A.) and examined under a fluorescence microscope (Nikon, Japan) with a confocal laser scanning system (Bio-Rad, Hercules, CA, U.S.A.).
Immunogold electron microscopy
ECV304 cells infected with O. tsutsugamushi were grown for seven days and were fixed with 0.5% glutaraldehyde, 4% paraformaldehyde and 3.5% sucrose and postfixed with 1% osmium tetroxide (OsO4). After dehydration in an ethanol series, the pellets were embedded in Epon 812. After the collection of ultrathin sections on Formvar/carbon-coated nickel grids, the grids were floated on drops of 3% sodium metaperiodate for 30 min. The immunogold labeling procedure (19) was done using MAb FS15 and 15 nm gold-conjugated goat anti-mouse IgG. After immunogold labeling, the grids were stained with uranyl acetate and lead citrate, and was viewed with a Hitachi H7100 electron microscope.
Flow cytometry assay and data processing
To quantify O. tsutsugamushi in ECV304 cells, infected cells were trypsinized, washed with PBS, and centrifuged at 300×g for 5 min. The cells were fixed with 70% ethanol for 1 hr on ice and stored at -20℃ until analyzed. Before their application to flow cytometry, fixed cells were washed with PBS and the resulting cell pellets were resuspended in 500 µL of FITC-conjugated MAb FS15 diluted with PBS, and analyzed with a flow cytometer (Becton Dickinson, Mansfield, MA, U.S.A.) equipped with a 15 mW argon laser. Data were collected on 10,000 cells and analysis was performed using LYSIS II software (Becton Dickinson). To calculate an estimate of the total mass of bacteria in the cells, the percentage of positive cells was multiplied by the mean fluorescence intensity of these cells to obtain the growth index (GI). The MIC was defined as the concentration of antibiotics at which GI was reduced by 90% relative to that of control cells that were infected and incubated in the absence of doxycycline.
RESULTS
Reactivity of MAb selected for the application in flow cytometry
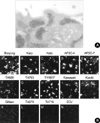
The tested MAbs could stain O. tsutsugamushi brightly in IFA (Fig. 1A). Although the MAbs showed similar morphologies of bacteria, their staining patterns differed slightly according to the target antigens. In addition, the bacterial particles stained by MAbs (FS10, FS15, and Rb167) directed against the 56-kDa scrub typhus antigen (Sta56) were brighter and bigger than those stained by MAbs (Rb134, M686-8, and Shim107) directed against other antigens. Therefore, we reasoned that MAbs against Sta56 are suitable as the primary antibodies for the use in flow cytometry. Among them, we selected FS15 because the target epitope of this MAb has been well characterized in a previous study (17). Furthermore, the antigenic epitope is located at the surface of the cell envelope as shown by immunogold electron microscopy (Fig. 2A). As Sta56 has strain-specific and group-specific epitopes, we tested the reactivity of FS15 to other strains of O. tsutsugamushi. As shown in Fig. 2B. MAb FS15 stained most of the O. tsutsugamushi strains (Boryong, Karp, Kato, AFSC-4, AFSC-7, TA686, TA763, TH1817, Kawasaki and Kuroki) originating from several Asian nations in IFA, except Gilliam, TA678, and TA716 strains.
Growth of O. tsutsugamushi measured by flow cytometry
To test the applicability of FITC-conjugated FS15 to the quantification of the total mass of O. tsutsugamushi by flow cytometry, we first used this MAb to determine the growth of O. tsutsugamushi. We infected the ECV304 cells with high (84.6% cells are infected 3-day postinfection) and low numbers (12.2% cells are infected 3-day postinfection) of O. tsutsugamushi and measured bacterial growth for seven days using flow cytometry. Under both conditions, flow cytometry demonstrated the significant growth of O. tsutsugamushi, which appeared at as early as one day after infection and progressed further until seven days after infection. This is evident in the right-hand shift of the entire peak of cells infected with high numbers of bacteria (Fig. 3A) and the appearance of a new peak in cells infected with low numbers of bacteria (Fig. 3B). Because three days incubation was sufficient to demonstrate bacterial growth by flow cytometric parameters, we selected this time point for subsequent work.
Determination of antibiotic susceptibility of O. tsutsugamushi by flow cytometry
To test the applicability of flow cytometry to the determination of the doxycycline susceptibility of the Boryong strain, which is the most prevalent strain in Korea (15), infected ECV304 cells were treated with various concentrations of doxycycline for three days, and then bacterial growth was measured by flow cytometry. As shown in Fig. 4A, the fluorescence peak shifted to left as the concentration of doxycycline increased, as indicated by the percentage of positive cells in each histogram. The percentage of positive cells infected with Boryong after treatment with 0.1 µg/mL doxycycline was 1.7% (Fig. 4A). In the presence of 0.1 or 0.2 µg/mL doxycycline, no bacterial growth was observed and the percentage of positive cells decreased to that of the control (uninfected) cells. Thus, the MIC was determined to be 0.1 µg/mL.
Next, we used our method to determine the doxycycline susceptibility of O. tsutsugamushi strain AFSC-4, which is reported to be resistant to doxycycline (5). The percentage of positive cells infected with AFSC-4 after treatment with 0.1 µg/mL doxycycline was 3.1% and the MIC was 0.1 µg/mL (Fig. 4B). When we compared the doxycycline susceptibility of the Boryong and AFSC-4 strains, both strains showed similar patterns of inhibition by doxycycline. The percentages of GI of Boryong and AFSC-4 strains after the treatment with 0.1 µg/mL doxycycline were 1.29% and 4.11% respectively, where 100% represented the percentage of infected cells without doxycycline treatment (Fig. 4C).
DISCUSSION
In this study, we have demonstrated that flow cytometry using specific MAb can be used to reliably quantify the total mass of O. tsutsugamushi. Traditional methods of quantifying other Rickettsia, such as the determination of the reduction in plaque formation or the dye-uptake assay (20), are seldom used for Orientia because it requires a long incubation time to form plaques (21). The most frequently used method involves counting Giemsa-stained bacteria after 3-4 days of antibiotic treatment (5). Whereas this method requires few special reagents, it is time-consuming and complicated owing to difficulties in quantifying the bacterial particles microscopically because of their tendency to form aggregates in cells (22, 23). With our method using flow cytometry, a right-hand shift of the entire peak or the appearance of a new peak was observed as O. tsutsugamushi divided. These results were in contrast to a previous trial using flow cytometry, in which only a slight right-hand shift occurred in the fluorescence peak of infected cells and seemed to be essentially qualitative rather than quantitative (14).
The MAb is the most important parameter in the sensitive detection of O. tsutsugamushi by flow cytometry. For the detection of bacteria in cells by flow cytometry, a primary antibody must have several properties: 1) it must be species-specific to detect various strains of O. tsutsugamushi, which is particularly important when we consider the antigenic heterogeneity of this bacterium; 2) target antigen should be expressed in large amounts, so that the antibody stains bacteria brightly; and 3) it must be available in large amounts so that the same reagent can be used in future applications.
We used a mouse MAb that is reactive against Sta56 of O. tsutsugamushi. This protein is abundantly expressed at the surface of the bacterium and is believed to be important in its pathogenesis (1). It has strain-specific and species-specific epitopes and is a major target of strain-specific immunity to O. tsutsugamushi. MAb FS15 was well characterized in our previous study as being reactive against amino acid residues 187-214 of Sta56. MAb FS15 was also reactive strongly on IFA against various strains of O. tsutsugamushi, including the most prevalent strain in Korea (15). Therefore, it can be used to test the antibiotic susceptibility of most strains. The morphologies of the bacteria stained with various MAbs on IFA differed according to the target antigen of each MAb. Interestingly, MAbs directed against Sta56 showed bigger and brighter bacterial particles than those detected by MAbs against other antigens of O. tsutsugamushi (Fig. 1). This is one of the desirable properties of an antibody to be used with flow cytometry. Our explanation for this finding is that Sta56 is an abundantly expressed protein at the cell surface of this bacterium. Of the MAbs directed against Sta56, we chose FS15 because it shows minimal nonspecific staining of host cells (data not shown).
The MIC of doxycycline was 0.1 µg/mL for both the Boryong and AFSC-4 strains when we measured bacterial growth after treatment with doxycycline for three days. This result is consistent with that of our previous study, in which we determined the antibiotic susceptibility of both strains using IFA (7). However, strain AFSC-4 is reported to be doxycycline-resistant and its MIC was reported to be 0.25 µg/mL (5). Although our approach produced an unexpected result, we would like to point that our method using flow cytometer is more objective than the microscopic count of Giemsastained bacterial particles used by authors of report for AFSC-4 strain (5). Although clinical experiences support the occurrence of infections by strains that are poorly responsive to doxycycline in northern Thailand, there are few reports about the biology of resistant strains and their mechanism for resistance. Therefore, further studies are required to clarify the mechanisms of doxycycline resistance of O. tsutsugamushi and the possible emergence of doxycycline-resistant in Korea.
In summary, we have improved the flow cytometric technique for the quantification of O. tsutsugamushi growth using specific MAb directed against Sta56. Our approach is sensitive in measuring the growing bacteria inside the cells. With this method, we measured the doxycycline susceptibility of the Boryong and AFSC-4 strains.


 XML Download
XML Download