

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Cyclooxygenase (COX) is a bifunctional enzyme with cyclooxygenase and peroxidase activities. The cyclooxygenase activity of COX is important for the initial conversion of arachidonic acid to prostaglandin (PG) G2, and the peroxidase activity of COX then converts it to PG H2. Finally, PG H2 is converted to other PGs by a variety of tissue specific enzymes. These PGs then exert their biologic actions on these tissues (1, 2). There are two isoforms of COX, a constitutive isoform COX-1 and an inducible isoform COX-2. COX-1 is constitutively expressed to fulfill its beneficial housekeeping roles. In contrast, COX-2 is frequently undetectable at baseline in most normal tissues, but it is rapidly induced by cytokines, tumor promoters, growth factors and carcinogens (3-5). Increased COX-2 expression has been demonstrated in various human cancers including gastric cancer and has been implicated in carcinogenesis (6-8). Furthermore, selective COX-2 inhibitors have been shown to have antineoplastic activities in various cancers (9, 10).
The several mechanisms by which COX-2 may contribute to carcinogenesis are beginning to unravel. These include that COX-2 stimulates tumor cell proliferation, inhibits apoptosis, increases the invasiveness of malignant cells, and enhance angiogenesis through the production of angiogenic factors (11-14). We previously reported that COX-2 may play a critical role in carcinogenesis by stimulating tumor angiogenesis in concert with vascular endothelial growth factor (VEGF) in human gastric cancer (15).
Regarding tumor cell proliferation, it is widely accepted that proliferative capacity may influence the clinical course, and hence patient prognosis. Ki-67, a nuclear antigen is expressed in all stages of the cell cycle except G0 and early G1, and it is often used to indicate the proliferative activity of tumors (16).
Tumor development, growth and progression are characterized by uncontrolled cell proliferation. This is usually the result of multiple genetic and epigenetic insults to the cell, particularily involving oncogenes and tumor suppressor genes (17). The p53, tumor suppressor gene is one of the most frequently mutated genes in human cancers. The p53 tumor suppressor gene is believed to play a pivotal role in preventing the uncontrolled cell proliferation characteristic of cancer. Recent studies reported that mutation of p53 may contribute to the increased COX-2 expression that is observed in malignant tissues (18, 19).
However, the role of increased COX-2 expression and p53 mutation in gastric cancer cell proliferation has not been fully established. The aim of this study was to evaluate the expression of COX-2 and p53 in gastric cancer and to examine the relationship between their expression and various clinicopathological characteristics including tumor cell proliferation.
MATERIALS AND METHODS
Sample selection
This study was based on an analysis of formalin-fixed, paraffin-embedded tissue specimens obtained from 119 patients who had gastric cancer and who underwent surgical resection at Chonnam National University Hospital from July 1994 to June 1995. None of the patients had received preoperative irradiation or chemotherapy before undergoing surgery. The specimens were taken from representative cancerous lesions over their greatest length and included adjacent noncancerous areas. Clinicopathological characteristics and survival data were obtained by hospital records, pathologist and physician contact when necessary. The tumors were staged at the time of surgery by the standard criteria for TNM staging using the American Joint Committee on Cancer (AJCC) (20). The mean age was 58.7±10.9 yr (mean±SD) with a range from 28 to 79 yr. Eighty-four patients were male, and 35 were female. The mean size of tumor was 5.1±2.7 cm (mean±SD) with a range from 0.5 to 15.0 cm. The mean follow-up period was 65.4 months with a range from 1.3 to 119.8 months.
Immunohistochemistry
All procedures for immunohistochemical staining were done by the Micro-Probe staining system (Fisher Scientific, Pittsburgh, PA, U.S.A.) based on capillary action (21). Formalin-fixed, paraffin-embedded tissue blocks were cut to 4-µm-thick sections for immunohistochemical staining. A standard avidin-biotin peroxidase complex method was used. Sections were deparaffinized using xylene and transferred to alcohol. Endogenous peroxidase activity was blocked using the 0.6% hydrogen peroxide and incubated for 5 min. Antigen retrieval was performed by microwave for 7 min. A monoclonal mouse immunoglobulin antibody to COX-2 (160112; diluted 1: 250; Cayman Chemical Co, Ann Arbor, MI, U.S.A.), p53 (DO-7; diluted 1:100; Dakopatts, Glostrup, Denmark), and Ki-67 (MIB-1; diluted 1:150; Dakopatts, Glostrup, Denmark) were used as primary antibodies. The primary antibodies, in the aforementioned concentrations were diluted in phosphate-buffered saline supplemented with 5% normal horse serum and 1% bovine serum albumin and then incubated with tissues for 25, 15 min at 45℃, and 90 min at room temperature, respectively for COX-2, p53 and Ki-67. Anti-mouse immunoglobulin G (Sigma, St. Louis, MO, U.S.A.) labeled with biotin was used as a secondary antibody for the detection of primary antibodies and slides were incubated for 10 min at 45℃. After multiple rinses with universal buffer, the slides were incubated in streptavidin-horseradish peroxidase solution (Biomeda, Foster, CA, U.S.A.) for 10 min. As the final step, the slides were developed for 10 min with the enzyme substrate, 3 amino-9-ethyl carbazole (AEC, Sigma, St. Louis, MO, U.S.A.). The slides were then counterstained with hematoxylin solution for 1 min (Research Genetics, Huntsville, AL, U.S.A.). After dehydration, the tissue was sealed with a universal mount (Research Genetics). For negative controls, the primary antibody was omitted and replaced with phosphate-buffered saline.
Interpretation of immunohistochemical stains for COX-2, p53 and Ki-67
The immunohistochemical staining was evaluated independently by two pathologists without knowledge of the clinical outcomes, analysing the intensity, area and pattern of immunohistochemical staining. In case of disagreement, the slides were reviewed and a consensus view achieved. COX-2 immunoreactivity score was calculated as the product of staining intensity and staining area. The staining intensity was arbitrarily graded on a scale of four grades: 0, no staining of cancer cells; 1, weak staining; 2, moderate staining; 3, strong staining. The percentage of staining area was also graded on a scale with four grades: 0, none; 1, <10%; 2, 10-50%; 3, >50%. Theoretically, the overall scores could range from 0 to 9. The specimens with a score of more than 4 were regarded as positive expression, and those with a score ≤4 as negative expression. p53 immunoreactivity was assessed as being positive only when tumors exhibited intense nuclear staining, and immunoreactivity was categorized into 2 groups: negative expression (less than 10% positive tumor cells) and positive expression (at least 10% positive tumor cells). A distinct nuclear immunoreactivity for Ki-67 was considered positive. The Ki-67 labeling index (LI) was determined by observing 1000 cancer cell nuclei in areas of the section with highst labeling frequency, and the percentage of Ki-67-labeled nuclei was used for analysis (16).
Statistical analysis
The χ2-test and Fisher's exact test, where appropriate, were used to compare expression of the COX-2, p53 and Ki-67 LI subgroups with various clinicopathological variables. The correlation between COX-2 or p53 expression and Ki-67 LI was analyzed for statistical significance using Student's t test. Actuarial survival rates of patients were evaluated according to the Kaplan-Meier method and the differences were tested with a log-rank test. The statistical software program used was Statistical Package for the Social Sciences (SPSS/PC+12.0, Chicago, IL, U.S.A.). The value of significance was taken as p<0.05.
RESULTS
Expression of COX-2, p53, and Ki-67 in gastric cancer
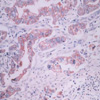
COX-2 immunoreactivity was cytoplasmic and was almost exclusively restricted to cancerous areas of pathologic specimens (Fig. 1). In some tumors, the COX-2 staining was diffuse, and in other tumors it was localized. In the normal gastric epithelia of non-cancerous area, no COX-2 immunoreactivity was observed. Immunoreactivity specific for p53 protein was evident in the nuclei of cancer cells and heterogenously distributed. Ki-67 immunoreactivity was almost found in the nuclei of cancer cells. Positive cells were frequent in the advancing margin of the tumor. Based on our criteria, expression of COX-2 and p53 in gastric cancer tissues, was demonstrated in 60.5% (72/119) and 34.5% (41/119), respectively.
Correlation between COX-2 or p53 expression and clinicopathological variables
The correlation between COX-2 or p53 expression and clinicopathological variables is shown in Table 1. Expression of COX-2 and p53 correlated significantly with depth of tumor invasion (p=0.005, 0.036 respectively). However, there was no association between COX-2 or p53 expression and tumor stage, status of lymph node, or distant metastasis. Furthermore, COX-2 and p53 expression did not associate with patient survival (p=0.927, 0.336 respectively) (Fig. 2A, B). The expression of p53 did not correlate with COX-2 expression (p=0.939) (Table 2).
Correlation between Ki-67 LI and clinicopathological variables
The Ki-67 LI for 119 tumors ranged from 7.6% to 85.8% with a mean Ki-67 LI of 49.5±15.5. When a mean Ki-67 LI value of 49.5 was chosen as the cut-off point for discrimination of the 119 patients into two subgroups, 57 patients were categorized as high Ki-67 LI and 62 as low Ki-67 LI. There was no significant difference in various clinicopathological variables including survival between the two subgroups (Table 1) (Fig. 2C).
Correlation between COX-2 or p53 expression and Ki-67 LI
The correlation between COX-2 or p53 expression and Ki-67 LI is shown in Table 3. The mean Ki-67 LI value of COX-2 positive tumors was 52.3±14.6 and significantly higher than that of COX-2 negative tumors (p=0.012). The mean Ki-67 LI value of p53 positive tumors was 50.7±14.7 and not significantly higher than that of p53 negative tumors (p=0.525). Combined analysis of COX-2 and p53 status showed that the mean Ki-67 LI value of both positive tumors was significantly higher than that of both negative tumors (p=0.025).
DISCUSSION
Most of antineoplastic effects of aspirin and other non-steroidal anti-inflammatory drugs have been linked to their ability to suppress PGs synthesis through inhibition of the activity of the inducible isoform COX-2 (9, 10). Overproduction of COX-2 and PGs has been found to accompany the development and progression of various human cancers including gastric cancer (6-8). However, the precise mechanisms by which they act is not yet understood. Therefore, understanding of underlying mechanisms of COX-2 action in cancer development and progression may lead to a better understanding of carcinogenesis. Recent studies have shown that COX-2 and PGs promote carcinogenesis as well as growth and spread of established tumors by stimulating cell proliferation, inhibiting apoptosis, increasing the invasiveness of malignant cells, and enhancing the production of VEGF, which promotes angiogenesis (11-14).
Ki-67 is recognized as a nuclear antigen present in proliferating cells but absent in resting quiescent cells. The Ki-67 LI, determined by Ki-67 immunohistochemistry, is a well-known proliferation marker and has been extensively used to estimate the growth fraction of tumors (16).
This study is the first to document the possible role of COX-2 expression for tumor cell proliferation in gastric cancer. In this study, the mean Ki-67 LI value of COX-2 positive tumors was significantly higher than that of COX-2 negative tumors. This result was concordant with result obtained by Yamagishi et al. (22). Also, Sawaoka et al. reported that both selective and non-selective COX-2 inhibitors exerted minimal effects on cell proliferation of human gastric cancer cell lines, which expressed lower levels of COX-2, but suppressed cell proliferation of human gastric cancer cell lines that overexpressed COX-2 (23). These results imply that COX-2 play a critical role in tumor cell proliferation of gastric cancer.
It is known that wild type p53, but not mutant p53, suppresses a variety of promoters that contain TATA elements (24). A recent in vitro study demonstrated that wild type p53 inhibits the formation of the complex between TATA binding protein and the promoter region of COX-2 gene in a cell-free system (18). Therefore, mutation of p53 may contribute to upregulate COX-2 expression, and it may be involved in the regulation of tumor cell proliferation.
The second aim of this study was to evaluate the correlation between p53 and COX-2 expression, with special reference to tumor cell proliferation. In this study, the expression of p53 did not correlate with COX-2 expression. In contrast, previous studies reported that tumors with p53 mutation were associated with higher level of COX-2 expression (18, 19). There are some possible explanations for this discrepancy. First, induction of COX-2 is not dependent on mutation of p53 alone, and regulated by many factors including cytokines, tumor promoters, growth factors, and oncogenes (3-5). Second, the mutant p53 protein is frequently more stable than wild type, and this stabilized protein product can therefore be detected using immunohistochemistry (25). However, the expression of p53 as revealed by immunohistochemistry might not indicate that the p53 is necessarily mutated and non-functional.
Cancer growth and progression is generally regarded as dependent on a high rate of cell proliferation rate and a low rate of apoptosis rate (17). Apoptosis and cell cycle arrest may be reduced when p53 is mutated, allowing cancer growth and progression (18, 19, 25). In this study, the relationship between status of p53 expression and Ki-67 LI was not statistically significant. This result may indicate that p53 mutation is predominantly associated with apoptosis, but not cell proliferation. Previous studies reported similar or contrary results (26, 27). The role of p53 mutation in tumor cell proliferation has yet been controversial. Combined analysis of p53 and COX-2 status showed that the mean Ki-67 LI value of both positive tumors was significantly higher than that of both negative tumors. This result suggests that molecular and biologic factors including tumor suppressor gene may act in the control of tumor cell proliferation.
The third aim of this study was to examine the expression of COX-2, p53, and Ki-67 in gastric cancer and their relationship with clinicopathological characteristics including patient prognosis. In this study, expression of COX-2 correlated significantly with depth of tumor invasion. However, there was no association between COX-2 expression and survival. There was some agreement, but also several differences between these results and other studies (8, 28, 29). We previously reported that COX-2 expression is associated with well differentiated and intestinal type pathway in gastric carcinogenesis and not useful for establishing prognoses for gastric cancer (8). It is well known that well differentiated and intestinal-type gastric cancer, which less invade and metastasize, are associated with a better prognosis than poorly differentiated and diffuse type cancer. Therefore, the prognostic significance of COX-2 is not established in gastric cancer. Also, expression of p53 correlated significantly with depth of tumor invasion. However, there was no association between p53 expression and survivial. Although relationship between p53 expression and poorer prognosis is suggested, it is still unclear whether p53 is an independent prognostic factor. Understanding of the molecular and biochemical mechanisms responsible for mutation of p53 is necessary before change of this molecule can be applied in clinical practice as a prognostic factor. The Ki-67 LI, an estabilished cell proliferation marker, often correlated to prognosis in gastric cancer (30, 31). However, in this study, when a mean Ki-67 LI value was chosen as the cut-off point for discrimination of the study patients into two subgroups as high Ki-67 LI and low Ki-67 LI, there was no significant difference in various clinicopathological variables including survival between the two subgroups. Liu et al. and Kanai et al. reported that the Ki-67 LI did not influence the prognosis in gastric cancer (32, 33). These controversial results may be related to the different score systems and different antibodies used. And because tumor growth and progression result from the imbalance between cell proliferation and apoptosis, alterations in the control of apoptosis can be as important as those of cell proliferation.
In conclusion, COX-2 expression is associated with tumor cell proliferation of gastric cancer. However, tumor cell proliferation through the regulation of COX-2 in gastric cancer may not be dependent on p53 status.




 XML Download
XML Download