

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Asthma is a chronic inflammatory disorder of airway with long history, even described by Hippocrates (1,2). Traditionally, asthma has been regarded as a clinical manifestation of allergic response to environmental agents (1). However, available epidemiological evidences suggest that the portion of asthma cases attributable to allergic sensitization (atopy) is less than one half (3). This indicates that the importance of etiological mechanism other than allergic mechanism might be neglected (3). This is especially true when considering the patients with severe asthma are more frequently nonallergic compared to those with mild-to-moderate asthma (4). The nonallergic asthma has been suggested as an autoimmune disease on the basis of frequent detection of circulating autoantibodies in patients with nonallergic asthma (1,5-7). However, a causal relationship between autoimmunity and asthma has not been demonstrated mainly due to lack of identification of pathogenetically and logically relevant autoantigen (8).
On a pathological viewpoint, airway epithelium has been suggested as a major target tissue for inflammatory response in asthma regardless of allergic sensitization (9) and could be a potential target of autoimmune response in asthma (10). We previously identified the cytokeratin 18 (CK18) protein as an airway epithelial autoantigen associated with nonallergic asthma while exploring the hypothesis that an autoimmune response to airway epithelial cell proteins might be involved in the pathogenesis of asthma (10).
Apoptosis and loss of adhesion of bronchial epithelial cells have been reported in patients with asthma and these have attributed to secretion of TNF-alpha and interferon gamma by T cells and eosinophils (11). Caspase-3 is a key mediator of apoptosis in mammalian cells and cleavage of CK18 protein by caspase-3 is a marker of early apoptosis in epithelial cells (12,13). It has been reported that the apoptotic caspase-3 protein was increased in bronchial epithelial cells of asthmatic patients, when compared with healthy controls (14).
In this study, the bindings of IgG autoantibodies to the fragments of CK18 protein cleaved by caspase-3 were analyzed by Western blot to investigate the antigen-binding characteristics of IgG autoantibodies in nonallergic asthma.
MATERIALS AND METHODS
Subjects
We used serum samples from three patients with nonallergic asthma who had circulating IgG autoantibodies against CK18 protein (Table 1). All patients had typical clinical history of asthma and documented reversibility of FEV1 greater than 15% after inhalation of bronchodilator. All patients underwent skin-prick test with 50 common aeroallergens (Bencard Co., Brentford, U.K.). Nonallergic asthma was defined when there was no positive skin reaction to any of the 50 common aeroallergens in the presence of a positive histamine control. All serum samples from subjects were aliquoted and stored at -20℃. All subjects gave informed consent, and the institutional review board approved this study.
Western blot analysis of IgG autoantibodies to CK18 protein
Recombinant human CK18 protein (Research Diagnostics Inc., Pleasant Hill Road Flanders, NJ, U.S.A.) was separated by discontinuous sodium dodecyl sulphate/polyacrylamide gel electrophoresis (SDS-PAGE) using an 8% resolving gel (pH 8.8) and a 4% stacking gel (pH 6.8). Following electrophoresis, proteins were transferred onto a polyvinylidine difluoride membrane (Bio-Rad Laboratories, Hercules, CA, U.S.A.). After the transfer, the membrane strips were probed with 1 mL of serum samples at dilution of 1 in 100 (v/v) for 2 hr at room temperature. After washes, the membrane was incubated with alkaline phosphatase-conjugated goat anti-human IgG (Sigma Chemical Co., St. Louis, MO, U.S.A.) for 2 hr at room temperature. After a final washing, the membrane was stained with a substrate solution (nitro blue tetrazolium/5-bromo-4-chloro-3-indoyl phosphate; Sigma Chemical Co.). A mouse monoclonal antibody to human CK18 protein (clone no. CY-90, Sigma Chemical Co.) known to recognize a epitope located in the region of 312-356 amino acids of CK18 was used as a positive control (15). Alkaline phosphatase-conjugated goat anti-mouse IgG (Sigma Chemical Co.) was used for detection of mouse monoclonal antibody.
In vitro cleavage of CK18 protein by caspase-3
1 µg of recombinant human cytokeratin 18 protein was incubated with 1 µg of caspase-3 (BD Biosciences, San Diego, CA, U.S.A.) in a 40 µL volume containing 50 mM HEPES, 0.1 M NaCl, 10% sucrose, 0.1% CHAPS, and 10 mM DTT for 4 hr at 37℃ and the reaction was stopped by adding SDS-PAGE sample buffer as described in a previous report (16).
RESULTS
Site-specific cleavage of CK18 protein by caspase-3
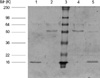
Caspase 3 has been reported to cleave 394DALD/S site of CK18 protein (16) (Fig. 1). We observed degradation of 49-kDa recombinant human cytokeratin 18 protein into 45-kDa fragment (corresponding N-terminal 1-397 amino acids; visible in Fig. 2) and 4-kDa fragment (corresponding C-terminal 398-430 amino acids; invisible in protein staining due to small molecular weight of this fragment in Fig. 2).
Binding of circulating IgG autoantibodies to fragments of CK18 produced by caspase-3
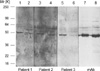
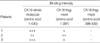
The binding capacity of IgG autoantibodies to N-terminal fragment (1-397 amino acid) of CK18 was maintained in one patient and reduced in other two patients (Fig. 3). IgG autoantibodies from all three patients did not show significant binding to C-terminal fragment (398-430 amino acids) of CK18 (Table 2). Mouse monoclonal IgG antibody to CK18 (clone CY-90; Sigma Chemical Co.) bound to the N-terminal fragment (1-397 amino acids) with same intensity of binding to whole molecule of CK18 protein (Fig. 3).
DISCUSSION
The presence of circulating IgG autoantibodies to CK18 could be just a reflection of epithelial damage in patients with nonallergic asthma considering recent evidence that the bronchial epithelium is more susceptible to injury in this group (17). Or the IgG autoantibodies could be a pathogenic factor contributing the development of chronic airway inflammation in nonallergic asthma.
Apoptosis is a biochemically and morphologically distinct form of programmed cell death, initiated by specific signals that activate specific caspases (13). As a result, the cell is eliminated by an intrinsic suicide program, resulting in DNA fragmentation, nuclear condensation, cytoskeletal reorganization, plasma membrane blebbing, and loss of cell adhesion (13). Apoptosis and loss of adhesion of bronchial epithelial cells have been reported in asthma and these have attributed to secretion of TNF-alpha and interferon gamma by T cells and eosinophils in asthmatic airway mucosa (11). Caspase inhibitor was shown to decrease airway inflammation in mouse model of asthma (18). Inhibition of epithelial cell apoptosis by corticosteroids has been suggested to be a mechanism of anti-asthmatic property of corticosteroids (19). Cleavage of CK18 protein by caspases and the exposure of specific CK18 neo-epitope identifiable by monoclonal antibody M30 (the same CK18 epitope exposed by caspase-3 in this study) has been reported to be a specific feature of early apoptosis (13). In previous study on idiopathic pulmonary fibrosis, the development of circulating IgG antibodies to cytokeratin 18 protein was suggested to be an secondary immune response to neo-epitopes of CK18 exposed by caspases during the apoptosis (20). In this study, we showed that IgG autoantibodies to cytokeratin 18 from patients with nonallergic asthma preferentially bound with whole molecule of CK18 protein, rather than the apoptotic fragments cleaved by caspase-3 and their antigen-binding characteristics were heterogeneous among the patients with nonallergic asthma. There is a certain limitation on the interpretation of these results due to the small number of patients (only three patients) because relatively large volumes of serum samples were needed for multiple repeated experiments. However, results of this study suggest that the autoantibody response to CK18 in nonallergic asthma might not be a secondary phenomenon associated with apoptotic death of airway epithelial cells due to chronic airway inflammation.
The idea of a possible involvement of autoimmunity in the pathogenesis of asthma has been proposed by earlier studies which demonstrated higher incidences of various autoantibodies against antigens in bronchial mucosa, paranasal sinus, lung, and endothelial cell in asthmatic patients compared to healthy controls (5-7). Analysis of T cell in airway fluid from patients with non-atopic asthma showed features of clonal expansion indicating the antigen-induced activation of T cells in the airway of non-atopic asthma (21). A significant correlation between T cell proliferative response to the 55-kDa endothelial autoantigen and the degree of airway obstruction has been reported in asthmatic patients (7). Patients with severe asthma can be clinically improved by anti-CD4 antibody treatment (22). In mouse model, non-atopic airway hyperresponsiveness could be passively transferred by CD4+ T cells (23). In rat model of autoimmune emphysema, transfer of pathogenic CD4+ T cells or antibodies developed by immunizing endothelial cell proteins caused emphysema (24). These suggest that a T cell response to an 'as-yet unidentified autoantigen' could be involved in the pathogenesis of nonallergic asthma (1,25). However, further studies including the T cell response to autoantigens and development of animal model for autoimmune asthma are necessary to determine the pathogenetic significance of autoimmunity in nonallergic asthma.


 XML Download
XML Download