

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Head and neck squamous cell carcinoma (HNSCC) which includes cancers of the larynx, pharynx, and oral cavity, show a wide range of geographical variations in incidence. Although tobacco and alcohol consumption are recognized as the major causes of HNSCC, geographic distribution of cancer incidence is not strongly correlated with areas of high tobacco and alcohol consumption. Also, only a small number of smokers actually develop HNSCC (1). This suggests a gene-environmental interaction where host components, likely genetics, have an influential role in the susceptibility to HNSCC (2). The presence of polymorphisms in genes coding for enzymes involved in tobacco-related carcinogen metabolism, such as the glutathione S-transferase (GST) and NAD(P)H:quinone oxidoreductase (NQO) enzymes, could explain this difference in susceptibility. It has recently been suggested that these inherited differences, in the capacity to metabolize such chemicals, are the modifiers of individual susceptibility to environmentally induced HNSCC (3).
Human GST enzymes can be subdivided into five main classes, alpha (A), mu (M), pi (P), theta (T), and zeta (Z). Each class includes one or more isoenzymes with different, but sometimes overlapping, substrate specificity (4, 5). As a result of conjugation with glutathione, the potential carcinogens are eliminated and DNA, or other important biomolecules, are protected against damage or adduct formation. The GSTP1 gene encodes for the isoenzyme GSTP1-1. GSTP1-1 is a major enzyme in the detoxification of tobacco-related metabolic products and, quantitatively, the most abundant GST enzyme in the human head and neck area (6). The GSTP1-1 enzyme level has been extensively studied in relation to tobacco-related malignancies. Elevated tissue levels of GSTP1-1 enzyme have been found in stomach, colorectal, bladder, oral, pharynx, larynx, lung, skin, and breast tumors compared with normal tissues of matched controls (7). Ali-Osman et al. have described a polymorphism of GSTP1, resulting from a single base change at codon 105, that produces an enzyme with reduced capacity for detoxifying a number of carcinogens (8). Recently, there have been several reports describing a higher risk of developing HNSCC at pharyngeal locations in patients who are Val homozygous for the GSTP1 Ile105Val variant (9).
NQO1 is also a detoxification enzyme that plays a critical role in protecting cells against chemically induced oxidative stress, cytotoxicity, mutagenicity, and carcinogenicity (10). NQO1 protects cells from oxidative damage by preventing the generation of reactive oxygen species through the reduction of endogenous quinines, and by reducing certain environmental carcinogens, such as nitroaromatic compounds, heterocyclic amines, and possibly cigarette smoke condensate. Furthermore, NQO1 reduces the formation of DNA adducts induced by benzo[a]pyrene 3,6-quinone, one of the important carcinogenic polycyclic aromatic hydrocarbons identified in cigarette smoke (11). However, under specific circumstances, metabolism by NQO1 yields more active products that can actually produce reactive oxygen species or undergo rearrangement to generate alkylating species (12). Therefore, NQO1 exerts both beneficial and harmful effects, which vary depending on the substrate. Many epidemiological studies have been performed to examine NQO1 polymorphism in tobacco-related cancers. These have shown that the NQO1 variant allele can modify the risk of colorectal cancer, lung cancer, renal cell carcinoma, and basal cell carcinoma (13-16).
In the present study, we investigated GSTP1 and NQO1 polymorphisms, in a large series of Korean patients with HNSCC and in matched controls, to determine the association between genetic polymorphisms and HNSCC risk. We also assessed the association between the tobacco exposure of all patients and GSTP1 and NQO1 polymorphisms.
MATERIALS AND METHODS
Study subjects
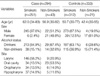
The study population consisted of 294 HNSCC cases and 333 cancer-free controls, recruited from the Department of Otolaryngology, at Asan Medical Center, from April 1994 to January 2004. All study participants were Korean. The case group consisted of 267 male patients (90.8%) and 27 female patients (9.2%); 251 cases were smokers with a mean age of 62 yr, and 43 were non-smokers with a mean age of 57 yr. Pathology reports were used to confirm the diagnosis of HNSCC for all cases. The control group consisted of 277 male patients (83.2%) and 56 female patients (16.8%); 312 were smokers with a mean age of 51 yr, and 21 were non-smokers with a mean age of 42 yr (Table 1). Controls were randomly selected from inpatients and outpatients with no history of cancer and were diagnosed with benign head and neck lesions. The distribution of primary cancer sites, among cases, is shown in Table 1.
For study purposes, subjects were divided into non-smokers and ever-smokers. Ever-smokers were defined as anyone who had smoked at least one pack-year; a pack-year was defined as the product of [{number of cigarettes consumed per day}×{duration of smoking (years)}]. Ever smokers were grouped as mild (≤30 pack-years) and heavy (>30 pack-years) smokers. Non-smokers were defined as individuals who had smoked less than one pack-year. Informed consent was obtained from all study subjects prior to participation.
Genotyping methods
After informed consent was obtained, each subject donated ten milliliters of blood. A B lymphocyte pellet, obtained from the buffy coat by centrifuging the whole blood, was used for DNA extraction using a DNA purification kit (GENTRA, Minneapolis, MN, U.S.A.). SNP genotyping was performed by SNP-IT™ assays using the SNPstream 25K™ System (Orchid Biosciences, NJ, U.S.A.). The genomic DNA region spanning the polymorphic site was PCR amplified using one phosphothiolated primer and one regular PCR primer (Table 2). The amplified PCR products were digested with exonuclease. The 5'phosphothiolates protected one strand of the PCR product from exonuclease digestion, resulting in the generation of a single-stranded PCR template. The single-stranded PCR template was then overlaid onto a 384 well plate that contained covalently attached SNP-IT™ primer extension, designed to hybridize immediately adjacent to the polymorphic site. Primer extension reaction was performed with the SNaPshot ddNTP Primer Extension Kit (Applied Biosystems, Foster City, CA, U.S.A.). To purify the product of the primer extension reaction, one unit of SAP (shrimp alkaline phosphatase) was added to the reaction mixture and was incubated at 37℃ for one hour, followed by fifteen minutes at 72℃ for enzyme inactivation. The DNA samples, containing the extension products and Genescan 120 Liz size standard solution, were added to Hi-Di formamide (Applied Biosystems). The mixture was incubated at 95℃ for five minutes, then placed on ice for another five minutes. It was then analyzed by electrophoresis in an ABI Prism 3700 Genetic analyzer. The results were analyzed using GeneScan and Genotyper software (Applied Biosystems).
Statistical analysis
The distributions of the GSTP1 and NQO1 genotypes between patients and controls were compared using the Mantel-Haenszel χ2 test. The frequencies of genotypes were analyzed between the cases and the controls within three subgroups of wild type, heterozygote, and homozygote mutant. Throughout this study, a p-value of <0.05 was considered statistically significant. Mantel-Haenszel χ2 test was also used to evaluate the association between the GSTP1 and NQO1 genotypes and smoking status in HNSCC risk. The association between genotypes and smoking status was evaluated within subgroups of the GSTP1 and NQO1 genotypes, the wild type group and the combined heterozygote/homozygote mutant group.
RESULTS
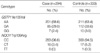
Table 3 shows the GSTP1 and NQO1 genotype distributions among cases and controls. The distribution of the GSTP1 and NQO1 genotypes among controls were in agreement with the Hardy-Weinberg equilibrium. The frequencies of GSTP1 and NQO1 genotypes in controls were similar to those previously observed in Koreans (17). When comparing the genotype distribution between control individuals and HNSCC patients, we observed that the frequency of the wild type GSTP1 and NQO1 genotype was slightly elevated in HNSCC patients, but the difference was not statistically significant (Table 3).
GSTP1 and NQO1 polymorphisms were associated with cigarette smoking and the risk of HNSCC in the case group (p<0.05). The frequency of genotype AA in GSTP1 Ile105Val was 83.7% in non-smokers, 70.1% in mild smokers, and 66.7% in heavy smokers; the frequency of the G allele containing genotypes (genotypes GA and GG) was 16.3%, 29.9%, and 33.3%, respectively. Genotype distributions were statistically significant (p=0.0114), and we observed that the GSTP1 105Val allele was associated with a tobacco dose-dependent increase in risk of HNSCC (Table 4). The frequency of genotype CC in NQO1 Trp139Arg was 100% in non-smokers, 98.7% in mild smokers, and 94.8% in heavy smokers, and the frequency of T allele containing genotypes (genotype TC and TT) was 0%, 1.3%, and 5.2%, respectively (Table 5). The NQO1 139Arg allele was associated with a tobacco dose-dependent increase in risk of HNSCC and these genotype distributions were statistically significant (p=0.0454). In the control groups, no significant difference of genotype distribution was found between wild type and mutant genotypes (Table 4, 5).
DISCUSSION
In this hospital-based, case-control study of 627 Korean patients, polymorphisms in two of the genes involved with metabolism of tobacco-induced carcinogens, GSTP1 Ile105Val and NQO1 Trp139Arg, were examined for association with HNSCC risk. We found no significant difference among the genotype distribution of polymorphic GSTP1 and NQO1 genes between HNSCC cases and controls. Although there are a large number of reports which suggest that polymorphisms of GSTP1 and NQO1 genes influence the susceptibility to cancer, the results of this present study suggest that the polymorphisms of the genes alone have only minor importance in the susceptibility to HNSCC. However, this finding is likely not due to the lack of a relationship between genetic polymorphisms and risk of HNSCC, but rather because biomolecular studies for genetic polymorphisms are still in the early phases of development.
To date, the role of several different DNA polymorphisms in detoxification genes have been examined in numerous case-control HNSCC studies. Results have often been conflicting, and some report weak-to-moderate associations, while others show no elevation of risk. These conflicting results may be due to relatively small samples sizes, or the lack of proper controls for other environmental exposures that are also risk factors for HNSCC. This study is also limited by the relatively small number of cases and controls. And the control population in this study was a convenience sample, and cases and controls were not balanced for confounding factors, age and sex. Therefore further studies with a larger number of subjects and well controlled risk factors such as age, sex, smoking, and alcohol exposures are needed.
In addition, the concept of genetic susceptibility is extremely complex and involves multiple cellular systems regulated by hundreds of genes (18), therefore, it is difficult to explain the association between genetic polymorphisms and risk of HNSCC by analyzing only one or two genes. In conclusion, a variety of analysis is needed, including a combined analysis of multiple polymorphic genotypes in detoxification enzymes, and of haplotypes to multiple enzymes associated with metabolisms of tobacco-induced carcinogens.
In our analysis, GSTP1 and NQO1 polymorphisms were associated with the status of cigarette smoking and, therefore an increased risk of HNSCC in the case group (p<0.05). For evaluation of smoking and genetic polymorphisms, we divide the subjects into three subgroups: non-smokers, mild smokers, and heavy smokers. Frequencies of the wild type GSTP1 and NQO1 genotypes were high in all three subgroups compared to frequencies of the mutant allele containing genotypes. Both GSTP1 and NQO1 genotype distributions in all three subgroups of the cases were statistically significant, and the GSTP1 105Val allele and NQO1 139Arg allele were associated with a tobacco dose-dependent increase in risk of HNSCC. It is likely that because GSTP1 and NQO1 are involved in the metabolism of specific tobacco smoke intermediates, ever smokers with the variant alleles, which are associated with reduced enzymatic activities, experience deleterious effects from reduced carcinogen detoxification. Therefore, there is a greater impact on HNSCC risk in smokers based on pack-years of smoking. This suggests that a predisposition to genetic factors, such as GSTP1 and NQO1 variants, plays a role in the development of HNSCC in smokers. However, there were no significant differences in the distribution of genetic variants among the smoking status in controls. There was also no significant difference in the genotype distribution of polymorphic GSTP1 and NQO1 genes between HNSCC cases and controls. From this point of view, environmental effects such as cigarette smoking are likely to dominate over genetic effects.
Several studies have examined the relationship between the GSTP1 polymorphisms and smoking status in HNSCC, and the results are not consistent. Ophuis et al. (19) found that HNSCC patients with the GSTP1 G allele containing genotypes (GA and GG) showed increased odds ratios for smoking compared with the wild type GSTP1 genotype. However, McWilliams et al. (20) found no association between the GSTP1 genotypes and smoking status in HNSCC risk. There are no studies performed to determine the relationship between the NQO1 polymorphisms and smoking in HNSCC. However, in bladder cancer, which is similar to HNSCC epidemiology in that it is strongly related to tobacco exposure, there are several studies that suggest the association between NQO1 genetic polymorphisms and the development of smoking related bladder cancer (21, 22).
Assessment of smoking or alcohol consumption is difficult, especially for the total accumulative exposure. Exposure usually takes place over many years and is not always consistent. In addition, different cigarette brands may yield completely different nicotine and tar exposures. Therefore, development of biomarkers, that could indicate the level of exposure and the period of consumption, would be more helpful.
In summary, to study the effect of different genotypes on the risk of developing HNSCC, we conducted a case-control study of two genetic polymorphisms in HNSCC. We were unable to demonstrate a significant association between genetic polymorphisms and risk of HNSCC. However, in analysis of the association between smoking and genetic polymorphisms, GSTP1 105Val allele and NQO1 139Arg allele were associated with tobacco dose-dependent increase in risk of HNSCC in the case group. Our findings suggest that the GSTP1 and NQO1 genotype polymorphisms may play an important role in the development of HNSCC, and that the association between cigarette smoking and HNSCC may be modulated by these genetic polymorphisms.



 XML Download
XML Download