

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The ovarian follicles have both epithelial and stromal layers in which cell migration or movement, cell division, specialization, differentiation, and death occur. A fluid-filled antrum develops and, at ovulation, the epithelial cells undergo an epithelial to mesenchymal transition into luteal cells. Human follicular fluid (HFF) contains a variety of biologically active products known to affect follicle growth and oocyte fertilization in the mammalian reproductive process. HFF also contains proteins produced by both granulosa and thecal cells in the ovary which may play essential roles in the regulation of follicular maturation. Some of these could also constitute markers of oocyte maturation during follicular development (1). For example, macrophage inflammatory proteins-3α, follistatin, activin, and TGFβ superfamily members correlating with oocyte maturation and folliculogenesis (2, 3) have previously been reported. HFF is related to endometriosis, an altered hypothalamic-pituitary-ovarian axis, spermatozoa-zona binding disease caused by cytokines, proteins similar to hormones (4, 5). HFF is a body fluid which contains inorganic salts, carbohydrates, mucopolysaccharides, lipids, proteins, steroids, peptide hormones, and growth factors within the ovarian follicle (5).
In combination with protein identification by mass spectrometry (MS), two-dimensional polyacrylamide gel electrophoresis (2-DE) has become an important tool for the study of proteomics (6-8). 2-DE gels use separated proteins in the first dimension by isoelectric focusing (IEF) in accordance with their charges and in the second dimension in accordance with their molecular weights (9). Here, we tried to identify new proteins in HFF by 2-DE with MS and make a standard 2-DE map of HFF from mature human follicles.
MATERIALS AND METHODS
Sample preparation
After informed consent was obtained, HFF was collected from five women (average of age 35 yr) of infertile couples with male factor undergoing in vitro fertilization at the In Vitro Fertilization (IVF) Department of CHA General Hospital, College of Medicine, Pochon CHA University (Seoul, Korea). When controlled ovarian hyperstimulation had resulted in the development of at least two follicles >18 mm in diameter, 10,000 IU of HCG (Profasi; Serono, Seoul, Korea) were administered. Oocyte retrieval was performed 35 hr later. Standard IVF and embryo transfer procedures followed. The different HFF aliquots were examined to detect cumulus-oocyte complexes. HFF was obtained and centrifuged at 13,000 rpm for 30 min to exclude granulosa cells and blood from the follicular fluid. Supernatants were also collected.
Two-Dimensional electrophoresis
For IEF, 50 µL of follicular fluid were mixed with 250 µL of rehydration solution (8 M urea, 100 mM DTT, 4% CHAPS, 0.5% carrier ampholytes, 40 mM Tris, 0.002% bromophenol blue dye) and immobiline 3-10 linear DryStrips (Amersham Bioscience, Piscataway, NJ, U.S.A.) were rehydrated together in a reswelling tray overnight. The protein sample solution was applied on immobilized pH 3-10 linear strips using an IPGphor system (Amersham Biosciences, Uppsala, Sweden). Focusing was performed in 3 steps (500 V for 1 hr, 1,000 V for 1 hr, and 8,000 V for 8 hr). Before the second dimension, strips were equilibrated for 15 min in SDS-PAGE equilibration buffer I (6 M urea, 0.375 M Tris, pH 8.8, 2% SDS, 20% glycerol, 130 mM DTT) and further incubated for 12 min in equilibration buffer II (6 M urea, 0.375 M Tris, pH 8.8, 2% SDS, 20% glycerol, 2.5% iodoacetamide). Equilibrated IPG strips were subjected to PAGE (6-20% gradient polyacrylamide gel) without stacking gels. The second-dimensional gels were run in the PROTEAN II xi 2DE Cells system (Bio-Rad, Hercules, CA, U.S.A.) at 10℃ for 5 hr at a constant voltage of 30 mA. For staining of gels with Coomassie brilliant blue (Bio-Rad), gels were equilibrated in a solution containing 50% (v/v) methanol, 5% acetic acid, and 25 g/L Coomassie brilliant blue R-250. Gels were rinsed in 30% (v/v) ethanol containing 7% acetic acid.
MALDI-TOF-MS
Protein spots were visualized with Coomassie Blue G-250. Stained gels were scanned using a BIO-RAD GS-800 scanner. Images were processed with Melanie 4 software. Spots targeted for digestion were excised (1×1 mm) and digested. Digested peptides were subsequently separated by mixture with the matrix for matrix assisted laser desorption/ionization (MALDI). Using ExPASy PeptIdent with an error tolerance of 100 ppm, information on the peptide mass fingerprinting data was searched against databases of relevant species.
RT-PCR
The expression of identified proteins in HFF were examined by RT-PCR using total RNA of granulosa cells that confirmed the results of 2-DE. RT-PCR was performed using the QIAGEN OneStep RT-PCR Kit (Qiagen, Valencia, CA) with the PCR DNA Thermal Cycler (Perkin Elmer, Norwalk, CT, U.S.A.). First-strand cDNA was synthesized from total RNA at 50℃ for 30 min and activated with HotStarTaq DNA polymerase by heating at 95℃ for 15 min. PCRs of first-strand cDNA were performed under the conditions of 1 min at 94℃, 40 sec at 56℃, and 1 min at 72℃. All primers in this study are listed in Table 1. The RT-PCR products were subjected to electrophoresis on 2% agarose gels.
RESULTS
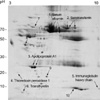
Isolated HFF samples were subsequently analyzed by high-resolution 2-DE using the IPG technique. A representative 2-DE of HFF is shown in Fig. 1. Coomassie blue-stained protein profile was obtained using 50 µL of follicular fluid and gel analysis by Melanie 4 software. 180 spots were resolved on the 3-10 pI strip. We attempted to analyze 40 protein spots that have a suitable amount for MALDI-time of flight (TOF) analysis. Ten spots were finally identified by peptide mass spectroscopy, and database searches against NCBInr and SwissProt were performed using the peptide mass fingerprint data listed in Table 1. The calculated protein molecular mass and pIs based on the amino acid sequence of each protein are also given. The peptide molecular ions (MH+) were obtained by MALDI-mass spectrometry (MS).
Identification of proteins from other 2-DE databases
The 2-DE pattern of HFF on pI 3-10 was similar to the silver-stained HFF 2-DE spots obtained by Anahory et al. using the 4-7 pI strip (10). We compared our data with the 2-DE gel protein profile published for human plasma and human follicular fluid (11). In the present study, our corresponding 2-DE spots were compared to the 6 spots found by Anahory et al. We noted, in addition, that a considerable degree of similarity existed between proteins in the follicular fluid and plasma/serum. Table 1 lists these proteins, and Fig. 1 is annotated to show the localization of the corresponding polypeptides on the Coomassie blue-stained 2-DE image. We were able to identify all of the proteins that were observed in the pH range 3-10 and had been previously reported, including: serum albumin (No. 1), serotransferin precursor (No. 2), apolipoprotein A-1 (No. 3), thioredoxin peroxidase 1 (No. 4), immunoglobulin heavy chain (No. 5), and transthyretin (No. 6).
Identification of new proteins in HFF
In addition to the above, we identified four new proteins which have not previously been reported: hormone sensitive lipase (HSL), apolipoprotein A-IV precursor, unnamed protein product 1 (UPP1), and unnamed protein product 1 (UPP2) (Table 1). Initially, these proteins were identified by peptide mass spectroscopy. Notably, HSL (No. 8) and apolipoprotein A-IV (apoA-IV) precursor (No. 9), which are related with lipid metabolism, were detected in HFF (Fig. 1). Using the BLAST program against the amino acid sequence database, homology searches revealed that UPP1 has homology with selenocysteine lyase, which is 100% homologous at the 194-445 amino acid sequences of selenocysteine lyase. Thus far, UPP2 has shown no homology with any protein by database search.
mRNA expression in granulosa cells
We examined whether 4 newly identified proteins originated from human granulosa cells. We isolated total RNA of human granulosa cells during IVF, and examined its mRNA expression (Fig. 2). Our semiquantitative RT-PCR showed that four newly identified proteins in HFF may originate from the human granulosa cell.
DISCUSSION
In the present study, we analyzed the protein expression patterns of several HFF samples by the direct rehydration method. The direct rehydration 2-DE method enabled the resolution of over 180 protein spots and led to the identification of new proteins. Our main goals were to provide a routine procedure for HFF sample preparation, to establish a standard 2-DE map of the normal HFF proteins, to monitor potential physiologic changes, and to take advantage of this approach to aid in the study of oocyte maturation and ovary-related disease diagnoses.
We found a total of 4 new proteins in HFF by 2-DE/MALDI-TOF MS (Table 1). UPP1 has been identified to human selenocystein lyase, which is a pyridoxal 5'-phosphate-dependent enzyme that specifically catalyzes the decomposition of L-selenocysteine to L-alanine and elemental selenium. Selenium is an essential trace element with several important biological functions. Several mechanisms have been proposed for the chemopreventive effects of selenocompounds (12-15). Therefore, UPP1 may have a potential role in regards to amino acid metabolism during oogenesis.
ApoA-IV was described as a component of intestine-derived, triglyceride-rich lipoproteins (16). Several studies have shown that the distribution of apoA-IV between lipoprotein-free and lipoprotein-associated forms can be altered in certain disorders of lipid metabolism, clearly indicating the metabolic role of this apolipoprotein. Moreover, there have been indications that apoA-IV may specifically be involved in reverse cholesterol transport, which suggests the importance of this apolipoprotein in cholesterol homeostasis and steroidogenesis (17).
HSL is a cytosolic neutral lipase that functions as the rate-limiting enzyme in the mobilization of free fatty acids in adipose tissue (18). HSL is an important enzyme in this protein kinase A (PKA)-activated process (19, 20), but the meager activation of HSL afforded by PKA in vitro, usually less than twofold, cannot account for the large increases (30-100-fold) in lipolysis observed upon elevation of PKA activity in mammalian adipocytes (21). The adipocyte protein content of β2-adrenoceptors, HSL, and the regulatory IIβ-component of PKA were found to be reduced by 70%, 55%, and 25%, respectively, in a common ovarian disorder, polycystic ovary syndrome (PCOS). Still, no change was observed in the amount of the catalytic subunit of PKA or β1-adrenoceptors. Thus, lipolytic catecholamine resistance by subcutaneous adipocytes in PCOS is probably due to a combination of reduced β2-adrenergic receptor levels, the regulatory IIβ-component of PKA, and HSL (22). HSL was detected within HFF in our study. Taking into consideration that HSL has been used as a marker of PCOS, this finding may be helpful for understanding the pathophysiology of PCOS. Recently, HSL expression was disrupted in a mouse model (23), and HSL was subsequently found to play a role in spermatogenesis, also acting as a lipase and a cholesterol esterase in various tissues (24). HSL and apolipoprotein are associated with lipid metabolism, thus these proteins are valuable in studies regarding PCOS and obesity.


 XML Download
XML Download