

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Androgens have a wide variety of biological effects, including several actions in hematopoiesis (1). The administration of testosterone was shown to increase hemoglobin levels in mice (2), and experiments with plethoric mice suggested that its major erythropoietic effect was exerted through the stimulation of erythropoietin (EPO) production by the kidney (3). This mechanism is unlikely to be involved in the beneficial effects of androgens in aplastic anemia patients, who produce abundant EPO; moreover, androgens have been shown to exert actions on hematopoietic cells independent of their effect on EPO secretion. In addition to increased hemoglobin levels, mice injected with testosterone derivatives had increased granulocyte and platelet numbers (4, 5). In vitro, androgens stimulated hematopoiesis in marrow suspension cultures (6-8) and enhanced both erythroid and myeloid colony formation in semisolid cultures (9, 10). It was demonstrated that testosterone was taken up by the nuclei of rat bone marrow cells and that it stimulated total RNA metabolism (11). Androgens have also been implicated in increasing the production of colony-stimulating factors by adherent cells from normal human blood (12). In addition, androgens have diverse, generally suppressive effects on the immune system (13, 14), which might be involved in their therapeutic effects in aplastic anemia.
In most of the previous studies regarding the in vitro effects of androgens on hematopoietic cells, whole bone marrow cells were used, rather than purified hematopoietic progenitor cells such as CD34+ cells. Furthermore, the question of whether androgens affect the survival of hematopoietic progenitor cells has not been well studied. In addition, the effects of androgens on the production of hematopoietic growth factors by bone marrow stromal cells (BMSCs), which might affect the growth and survival of hematopoietic progenitors, are poorly understood. With these questions in mind, we investigated the effects of androgens, such as testosteone, 5β-dihydrotestosterone (5-DHT), and oxymetholone, on the survival of purified normal hematopoietic progenitor cells and on the production of cytokines by normal BMSCs. Here, we show that direct survival-enhancing or growth-stimulatory effects of androgens on hematopoietic progenitor cells were minimal and mostly restricted to mature erythroid progenitors. Oxymetholone and 5-DHT stimulated the production of several cytokines in bone marrow stromal cells, and oxymetholonestimulated stromal cells better supported the survival of hematopoietic progenitor cells.
MATERIALS AND METHODS
Cells and cell lines
CD34+ cells were purified from bone marrow obtained from normal donors undergoing bone marrow harvest, after informed consent. Following Ficoll separation, CD34+ cells were isolated using the MACS system (Miltenyi Biotec, Auburn, CA, U.S.A.) according to the manufacturer's instructions. Only cell preparations with purity of 95% CD34+ or greater by flow cytometric analysis were used in the experiments. Primary human BMSCs were prepared by culturing CD34-negative cells (the cells that remained after purification of the CD34+ cells from the bone marrow mononuclear cells) in MEM-α (Gibco BRL Life Technologies, Grand Island, NY, U.S.A.) supplemented with 12.5% fetal bovine serum (FBS), 12.5% horse serum (Gibco), 10-6 M hydrocortisone (Sigma, St. Louis, MO, U.S.A.), 5×10-5 M 2-mercaptoethanol (Sigma), streptomycin, and penicillin (hereafter referred to as LTC medium). After confluence was reached, the adherent cells were harvested and maintained in LTC medium with weekly passage. After three passages, neither CD45+ cells nor PECAM-1+ cells were detected by flow cytometric analysis. The murine bone marrow stromal cell line MS-5 was grown in MEM-α supplemented with 10% FBS and was passaged weekly.
Hormones and cytokines
Testosterone, 5β-dihydrotestosterone (5-DHT), and β2-estadiol were purchased from Sigma. Oxymetholone was kindly provided by Hanseo Pharmaceutical Co. (Seoul, Korea). The hormones were formulated as 1×10-2 M stock solutions using ethanol as diluent and stored at -20℃. The following cytokines were used: interleukin-3 (IL-3), stem cell factor (SCF), flk-2 ligand (FL), thrombopoietin (TPO), EPO, and stromal cell-derived factor-1α (SDF-1α) (all purchased from R&D Systems, Minneapolis, MN, U.S.A.).
Flow cytometry
A total of 5×104 to 1×105 cells were incubated at 4℃ for 30 min with phycoerythrin (PE)-conjugated anti-CD34 monoclonal antibody (HPAC-2; Becton-Dickinson, San Jose, CA, U.S.A.) and were analyzed using a Coulter Elite flow cytometer (Coulter Electronics Ltd., Hialeah, FL, U.S.A.). Isotope-identical antibodies served as controls. To detect apoptosis, cells were stained with FITC-conjugated annexin V (BD PharMingen, San Diego, CA, U.S.A.) alone or together with propidium iodide (PI; BD PharMingen) according to the manufacturer's instructions and analyzed by flow cytometry.
Clonogenic assay in methylcellulose
Cells (1×103 CD34+ cells or the equivalent) were plated in triplicate in 35-mm tissue culture dishes containing 1 mL of assay medium consisting of Iscove's modified Dulbecco medium (IMDM; Gibco), 1.2% methylcellulose (Stem Cells, Vancouver, Canada), 30% FBS, 50 µM 2-mercaptoethanol, 2 mM L-glutamine (Gibco), and 0.5 mM hemin (Sigma), supplemented with 20 ng/mL SCF, 50 ng/mL IL-3, and 6 U/mL EPO. After 7-14 days of incubation at 37℃ in 5% CO2 in air, the numbers of colony-forming unit-granulocyte/macrophage (CFU-GM), burst-forming unit-erythroid (BFU-E), and colony-forming unit-erythroid (CFU-E) were scored under the inverted microscpoe.
Preparation of conditioned media (CM)
Conditioned media were prepared from MS-5 cells and primary human BMSCs grown in the appropriate medium in T25 culture flasks. When confluence was reached, the cultures were thoroughly rinsed, and 3 mL of serum-free X-VIVO medium (BioWhittaker, Walkersville, MA, U.S.A.) was added to each flask, with or without the inclusion of androgens. After a 72-hr incubation at 37℃, the supernatants were harvested, filtered through 2-micron membranes, and stored at 4℃ until analyzed.
Enzyme-linked immunosorbent assay (ELISA)
The concentrations of SDF-1α and SCF in the conditioned media were measured using commercial ELISA kits (R&D Systems) according to the manufacturer's instructions. The optical density was measured using a spectrophotometer (Molecular Device Co., Sunnyvale, CA, U.S.A.). The standard curves and calculated levels of the cytokines were obtained using the program Softmax™ (Molecular Device Co.).
Reverse transcriptase-polymerase chain reaction (RT-PCR)
Total RNA was prepared from primary human BMSCs using Trizol™ (Gibco) according to the manufacturer's instructions. After purification, 1 µg of RNA was reverse-transcribed using SuperScript reverse transcriptase (Gibco) and the universal primer oligo dT15 (Promega, Madison, WI, U.S.A.). One µL of the cDNA products was then added to 24 µL of PCR buffer (Gibco) supplemented with 2 mM MgCl2, 0.2 µM specific primers (Table 1), and 1 U/reaction tube of Koma Taq polymerase (Koma International, Seoul, Korea). The PCR cycles consisted of 1 min at 94℃, 45 sec at 55-65℃, and 1 min at 72℃; 30 cycles were carried out using a GeneAmp PCR system (Perkin Elmer, Norwalk, CT, U.S.A.).
Northern blot analysis
The expression of SDF-1 mRNA in primary human BMSCs was detected by Northern hybridization. Briefly, 20 µg of total RNA was loaded in each lane, separated on 1.2% agarose gels containing 2.4% formaldehyde, and then transferred onto nylon membranes (Schleicher & Shuell, Keene, NH, U.S.A.) for 24 hr by the capillary transfer method. All probes were radiolabeled using a random priming protocol (Amersham Life Science, Arlington Heights, IL, U.S.A.). The membranes were hybridized with random primer-labeled SDF-1 probe for 24 hr, washed, and then subjected to autoradiography.
Western blot analysis
Western blotting was used to detect the expression of SDF-1 protein in primary human BMSCs. Cells were pelleted, washed in phosphate-buffered saline, and lysed by the addition of SDS sample buffer composed of 62.5 mM Tris-HCl (pH 6.8), 6% (w/v) SDS, 30% glycerol, 125 mM DTT, and 0.03% (w/v) bromophenol blue. Total cell samples were sonicated, lysed, and denatured by boiling for 5 min. Equal amounts of protein from each sample were electrophoresed on 15% SDS-polyacrylamide gels and transferred to nitrocellulose membranes (Amersham). The membranes were blocked for 1 hr with TBS containing 5% (w/v) milk and 0.1% Tween, and then incubated with the primary mouse monoclonal antibody (12G5; R&D Systems) overnight at 4℃. The blots were washed with TBST buffer, incubated with anti-mouse secondary antibody (Amersham) for 2 hr, and developed using West-Zol™ Plus (iNtRON Biotechnology, Seoul, Korea).
RESULTS
Androgens stimulates clonal growth of CFU-E, but not BFU-E or CFU-GM
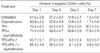
We examined the effect of the addition of 1×10-7 to 1×10-5 M oxymetholone or other hormones to normal BM CD34+ cells in a standard methylcellulose assay supplemented with the hematopoietic growth factors SCF, IL-3, and EPO. Oxymetholone did not influence the numbers of CFU-GM or BFU-E; however, there was a slight increase in the number of CFU-E in the presence of 1×10-7 or 1×10-6 M, but not 1×10-5 M, oxymetholone compared with controls grown without added hormones. The addition of testosterone, 5-DHT, or β-estradiol also induced an increased number of CFU-E, but did not affect the number of CFU-GM or BFU-E in this assay (Table 2).
Short-term treatment of CD34+ cells with androgens in the absence of HGFs enhances the survival of CFU-E, but not CFU-GM or BFU-E
Normal BM CD34+ cells were pre-incubated for 36 hr in the absence of serum and androgens before being plated in a standard methylcellulose clonogenic assay. The addition of 1×10-7 to 1×10-5 M oxymetholone during the pre-incubation period did not affect the formation of CFU-GM or BFU-E; however, 1×10-6 M oxymetholone increased the number of CFU-E compared with controls pre-incubated without added hormones. Similarly, pre-incubation with 5-DHT or testosterone induced an increase in the number of CFU-E. Unexpectedly, the addition of 5-DHT, testosterone, or β-estradiol caused a slight decrease in the number of CFU-GM (Table 3).
Testosteone and 5-DHT at a high concentration, but not oxymetholone, enhance apoptosis of normal BM CD34+ cells induced by growth factor deprivation
Normal BM CD34+ cells were incubated for 36 hr in serum- and hematopoietic growth factor (HGF)-free IMDM, with and without androgens at a concentration of 10-5 M. After the cells were stained with propidium iodide and annexin V, the level of apoptosis was analyzed using flow cytometry. The percentage of annexin-positive apoptotic cells was not changed by the addition of oxymetholone (51.5±5.1%) compared with controls (52.6±5.6%). Similar results were obtained at lower concentrations of oxymetholone (10-9 to 10-6 M; data not shown). Again unexpectedly, both testosterone and 5-DHT at this concentration significantly increased the percentage of annexin-positive cells (62.2±5.9%, p<0.05; 61.7±6.4%, p<0.05, respectively) (Fig. 1A). The addition of either SDF1 or SCF in combination with 5-DHT partially relieved the increase in apoptosis induced by 5-DHT, and the addition of both SDF-1 and SCF completely reversed it (Fig. 1B).
Oxymetholone does not affect cytokine-mediated alteration of survival of normal BM CD34+ cells
To further define the role of androgens in the survival of hematopoietic progenitor cells, we examined the time course of apoptosis. Normal BM CD34+ cells were incubated for 7 days in IMDM in the absence or presence of a combination of low concentrations of growth factors (5 ng/mL TPO+5 ng/mL FL+5 ng/mL SCF, hereafter referred to as TFS). This combination of growth factors was expected to enhance the survival, rather than the proliferation, of CD34+ cells. In the absence of all cytokines, annexin-positive cells markedly increased over time; the addition of oxymetholone did not delay the development of apoptosis. In the presence of TFS, the percentage of annexin-positive cells was much lower at every time point compared with that of cells incubated in the absence of TFS, regardless of CD34 positivity. Again, the addition of oxymetholone in combination with TFS did not result in any changes in the level of apoptosis (Table 4).
We next examined whether oxymetholone affects IFN-γ-mediated alterations in the clonal growth and survival of hematopoietic progenitor cells. In a standard methylcellulose clonogenic assay containing SCF, IL-3, and EPO, the addition of INF-γ caused a significant decrease in the numbers of both CFU-GM and BFU-E; the inclusion of 10-9 to 10-5 M oxymetholone had no effect on this INF-γ-induced inhibition of clonal growth (data not shown). Normal BM CD34+ cells were incubated for 7 days in IMDM in the absence or presence of TFS. The addition of IFN-γ in combination with TFS significantly decreased the percentage of annexin-negative CD34+ cells in a dose-dependent manner, while increasing the percentage of annexin-negative/CD34-negative cells and annexin-positive cells. Again, the inclusion of oxymetholone did not alter the effects of IFN-γ on the survival of CD34+ cells (Table 4).
5-DHT, but not testosterone or oxymetholone, down-regulates SDF-1 production in bone marrow stromal cells
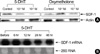
The possibility that androgens might be involved in the regulation of SDF-1 production in BMSCs was explored. Conditioned medium collected after a 72-hr incubation with untreated MS-5 cells contained a high concentration of SDF-1α (64.3±6.4 ng/mL). The addition of 5-DHT at the beginning of the culture period diminished the concentration of SDF-1 in the conditioned medium in a dose-dependent manner, whereas the addition of oxymetholone, testosterone, or β-estradiol did not (Table 5). Based on these observations, Western blot analysis was performed, using primary human BMSC lysates. Treatment of the BMSCs with 5-DHT definitely decreased the amount of SDF-1 in a dose-dependent manner, indicating that this hormone down-regulates the production of SDF-1 in the cells (Fig. 2A). Northern blot analysis revealed that the expression of SDF-1 mRNA in the BMSCs was also down-regulated by 5-DHT (Fig. 2B).
Androgens modulate the production of HGFs in BMSCs
The possibility that androgens might influence the production of HGFs in BMSCs was also investigated. Primary human BMSCs were incubated in serum-free medium for 24 hr with or without androgens, and the expression levels of the mRNAs for various HGFs were analyzed by RT-PCR. Oxymetholone and 5-DHT induced increased expression of the mRNAs of IL-7, SCF, and TPO; decreased expression of IL-6 mRNA; and no change in the expression of FL mRNA (Fig. 3). We also used ELISA assays to examine the levels of SCF in the conditioned media collected after a 72-hr incubation with primary human BMSCs in the presence or absence of the hormones. The addition of 5-DHT and oxymetholone at the beginning of the culture period increased the concentration of SCF in the conditioned medium in a dose-dependent manner, whereas the addition of testosterone or β-estradiol did not (Table 6).
BMSCs treated with oxymetholone better support the survival of hematopoietic progenitor cells
Normal BM CD34+ cells were incubated for 36 hr in conditioned media harvested from primary human BMSCs incubated in the presence or absence of 10-5 M oxymetholone. There was a lower percentage of annexin-positive apoptotic cells in the culture incubated in conditioned medium from oxymetholone-treated BMSCs than in the culture incubated in conditioned medium from untreated BMSCs (Table 7).
DISCUSSION
In the present study, we showed that androgens, including oxymetholone, have a modest growth- and survival-enhancing effect on CFU-E, but not on CFU-GM or BFU-E. These effects, which were restricted to mature erythroid progenitors, might be the basis for the observed androgen dependency among responders to androgen therapy for aplastic anemia (15). From the combined findings, it is clear that androgens do not exert their myelostimulatory effects at the level of hematopoietic stem cells or immature progenitor cells. It was quite unexpected that testosterone and 5-DHT at a high concentration enhanced to some extent the apoptosis of normal BM CD34+ cells induced by serum deprivation and growth factor deprivation, rather than decreased it. These results differ from observations in previous studies that found testosterone and its derivatives enhanced the survival of hematopoietic progenitor cells (8). This disparity could be attributable to the type of cells analyzed in the experiments. In the previous studies, bone marrow cells were used without purification and thus were a mixed population of hematopoietic progenitor cells and various accessory cells present in the BM. It is possible that the observations in the previous studies were the result of the modulation of survival by accessory cells, such as BMSCs, rather than the direct action of the hormones on the hematopoietic progenitor cells. To our knowledge, this is the first time that enhancement of apoptosis of normal BM CD34+ cells by testosterone and 5-DHT has been reported. This finding is less surprising than it initially might appear, because androgens have been shown to induce apoptosis and growth inhibition of non-lymphocytic leukemic cells (16, 17).
IFN-γ is well known as a hematopoiesis-inhibitory cytokine (18-20) and has been implicated in the pathogenesis of bone marrow failure (21, 22). In the present study, oxymetholone did not rescue colony-forming cells from IFN-γ induced inhibition of clonal growth of BM CD34+ cells in methylcellulose cultures. Furthermore, oxymetholone did not mitigate IFN-γ-induced suppression of growth factor-mediated survival of CD34+ cells. These results suggest that direct action of oxymetholone on hematopoietic progenitor cells is minimal, raising the possibility that the myelostimulatory effects of oxymetholone are attributable to other mechanisms.
Stromal cell-derived factor-1 (SDF-1), which is constitutively produced in BMSCs, plays a central role in bone marrow homing of hematopoietic stem cells by signaling via its receptor, CXCR4 (23). Recently, it has also been demonstrated that SDF-1 is involved in the positive regulation of hematopoiesis. SDF-1 enhances both the survival and proliferation of hematopoietic progenitors, especially in synergy with other hematopoietic growth factors (24-26), while inhibiting the cycling of very primitive hematopoietic cells, both in vitro and in vivo (27). Therefore, it is possible that the down-regulation of SDF-1 production in BMSCs would lead to the suppression of hematopoietic cell survival and growth. In the present study, we showed that 5-DHT, but not testosterone or oxymetholone, down-regulated SDF-1 production in BMSCs and that 5DHT-induced increases in apoptosis were nearly completely relieved by the addition of SCF and SDF-1. This paradoxical interaction between androgens and SDF-1 might be part of a regulatory mechanism for in vivo homeostasis of hematopoiesis. Androgens are known to suppress B lymphocyte development (28), and it has been demonstrated that this suppression is mediated through the up-regulation of the production of tumor growth factor-beta in BMSCs (29). In this context, it is possible that the down-regulation of SDF-1 production in BMSCs by 5-DHT contributes to the suppression of B lymphocyte development, because SDF-1 also plays a role in the development of B lymphocytes (30).
Androgen receptor is ubiquitously expressed in the bone marrow cells of both males and females, with the exception of lymphoid and erythroid cells (31). Testosterone derivatives down-regulated IL-6 production in BMSCs in vitro (32), which was consistent with the observation that orchiectomy increased bone marrow IL-6 levels in mice (33). However, little was known concerning whether testosterone or its derivatives are involved in regulating the production of hematopoietic growth factors. In the present study, we clearly showed that androgens modulate the mRNA expression of several hematopoietic growth factors in BMSCs. Oxymetholone up-regulated the production of SCF via transcriptional regulation, and conditioned medium derived from oxymetholone-treated BMSCs better supported the survival of BM CD34+ cells. These results could provide an explanation for the observed oxymetholone-mediated enhancement of granulopoiesis and thrombopoiesis, as well as erythropoiesis, in vivo.
In summary, androgens including oxymetholone directly stimulated the clonal growth and survival of mature erythroid progenitor cells, but not that of immature erythroid progenitors or granulocytic progenitors. Oxymetholone increased the capacity of BMSCs to support hematopoietic progenitor cells, most likely by up-regulating the production of a couple of hematopoietic growth factors, including SCF.
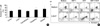







 XML Download
XML Download