

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Gabapentin is a novel antiepileptic agent and a structural analog to γ-aminobutyric acid (GABA) (1). Studies of the activity profile of gabapentin have shown that intrathecal gabapentin counteracts injury-induced hyperalgesia, but it does not reduce acute nociception (2-4). Although the specific mechanism of gabapentin's antinociception is not certain, N-methyl-D-aspartate (NMDA) and alpha-amino-3-hydroxy-5-methtyl-4-isoxazolepropionate (AMPA) receptors have been suggested as sites of action in the spinal cord (5).
Glutamate, the excitatory amino acid (EAA), is involved in the transmission of nociceptive information in the spinal cord (6). The excitatory effect of glutamate is considered to be mediated through at least two distinct classes of receptors, NMDA and AMPA receptors (7-9). Intrathecal NMDA antagonists have various effects on acute nociception, but they inhibit injury-induced hyperalgesia (10-15). On the other hand, intrathecal AMPA antagonists alleviate acute nociception with different effects on injury-induced hyperalgesia (2, 7, 16).
These observations suggest that spinal gabapentin, NMDA antagonists and AMPA antagonists may or may not be against nociception according to the nociceptive conditions. However, there has been little data regarding the interaction between gabapentin and antagonists for NMDA or AMPA receptors at the spinal level in the formalin test, which is a preclinical pain model showing acute nociception followed by injury-induced hyperalgesia.
The aim of this study was threefold: 1) to examine the effect of intrathecal gabapentin, NMDA antagonist (MK801) and AMPA antagonist (NBQX) in the rat formalin test, 2) to evaluate the consequence of gabapentin on the effect of MK801 or NBQX in acute nociception, and 3) to determine the spinally mediated antinociceptive interactions between gabapentin and MK801 or NBQX in the injury-induced hyperalgesia.
MATERIALS AND METHODS
This work was carried out with permission from the Animal Care Committee of our Research Institute of Medical Science.
Male Sprague-Dawley rats (250-300 g) were used. Rats were housed in group cages and kept in a vivarium, maintained at 22℃ with a 12 hr alternating night/day cycle, and were given water and food ad libitum. For drug administration, intrathecal catheters were advanced caudally 8.5 cm into the subarachnoid space through an incision in the atlantooccipital membrane during enflurane anesthesia (17).
Rats showing postoperative neurologic sequelae were immediately killed with an overdose of volatile anesthetics. After surgery, the rats were kept in individual cages and allowed to recover for 4-5 days.
The following drugs were used in this study: gabapentin (1-[aminomethyl] cyclohexanacetic acid; Sigma Chemical Co., St. Louis, MO, U.S.A.), MK801 (NMDA receptor antagonist; Research Biochemical Internationals [RBI], Natick, U.S.A.) and NBQX (AMPA receptor antagonist; Tocris Cookson Ltd., Bristol, U.K.). Gabapentin and MK801 were dissolved in normal saline. NBQX was dissolved in dimethylsulfoxide (DMSO). Intrathecal administration of these agents was performed using a hand-driven, gear-operated syringe pump. All drugs were given in a volume of 10 µL solution, followed by an additional 10 µL of normal saline to flush the catheter.
The formalin test was used as a nociceptive test (4). 50 µL of 5% formalin solution was injected subcutaneously into the plantar surface of the hindpaw. After formalin injection, rats show a characteristic pain behavior, biphasic flinching/shaking of the injected paw. Such pain behavior was therefore quantified. The interval from 0-9 and 10-60 min after the formalin injection was defined as phase 1 and phase 2 of the formalin test, respectively. Upon completion of the 60 min observation, the rats were immediately killed with an overdose of volatile anesthetics.
Animals were behaviorally tested four to five days after intrathecal catheterization. After acclimation for 15-20 min in a restraint cylinder, rats were then placed into one of the experimental groups. The drug vehicles (saline or DMSO) were used as a control for intrathecal drugs. The rats were only used once.
The first series of experiments were performed to examine the time course and dose-dependency of intrathecal gabapentin (10, 30, 100 and 300 µg), MK801 (3, 10 and 30 µg) and NBQX (0.3, 1 and 3 µg) in the formalin test. Intrathecal drugs were given 10 min before the formalin injection. Each ED50 value (effective dose producing a 50% reduction of control formalin response) of three agents was separately determined in two phases.
To determine the characteristics of interaction between gabapentin and the NMDA antagonist, or between gabapentin and the AMPA antagonist, a fixed dose analysis and an isobolographic analysis were used (18).
A fixed dose analysis was used in phase 1 because intrathecal gabapentin did not produce an antinociceptive effect during phase 1. A fixed dose of gabapentin (300 µg) was intrathecally coadministered with various doses of MK801 or NBQX. An isobolographic analysis was used to define the property of interaction during phase 2. This method is based on comparisons of doses that are determined to be equally effective. First, each ED50 value was determined from the dose-response. Then, the ED50 values of the mixture were calculated from the dose-response curves of the combined drugs and used for plotting the isobologram. In order to understand the magnitude of the interaction, a total fraction value was also calculated.
Total fraction value
=ED50 [(drug 1+drug 2)/drug 1+(drug 2+drug 1)/drug 2]
The fraction values indicate what portion of the single ED50 value was accounted for by the corresponding ED50 value for the combination. Values near 1 indicate an additive interaction, values greater than 1 imply an antagonistic interaction and values less than 1 indicate a synergistic interaction. The mixtures were delivered intrathecally 10 min before the formalin test.
To examine the behavioral changes by gabapentin, MK801 and NBQX, the highest dose of each drug was given intrathecally to the additional rats. Motor function was assessed by the placing-stepping reflex and the righting reflex (14). The first was evoked by drawing the dorsum of either hind paw across the edge of the table. Healthy rats generally try to put the paw ahead into a position for walking. The other was evaluated by placing the rat horizontally with its back on the table. Healthy rats give rise to an immediate and coordinated twisting of the body into an upright position.
Data are expressed with a mean±SEM. The time-response data are presented as the number of flinches. The dose-response data are presented as the total sum of flinches in each phase. To calculate the ED50 values of each drug, the number of flinches was converted into percentage of control according to the following formula:
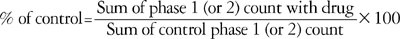
Dose-response data were analyzed by one-way analysis of variance (ANOVA) with Scheffe for post hoc. The dose-response lines were fitted using least-squares linear regression and ED50 and its 95% confidence intervals were calculated according to the method described by Tallarida and Murray (19).
The difference between theoretical ED50 and experimental ED50 was examined by t-test. A p value less than 0.05 was considered significant.
RESULTS
A biphasic flinching response of the injected paw was observed in all rats after subcutaneous injection of formalin into the hindpaw.
Fig. 1 displays the time course of intrathecal gabapentin, MK801 and NBQX in the formalin test. The sum of the number of flinches in the control group did not differ statistically in either phase (saline vs. DMSO: 20±2 vs. 20±1 in phase 1,160±8 vs. 158±13 in phase 2). Intrathecal MK801 and NBQX, but not gabapentin, produced a dose-dependent reduction of flinching response during phase 1 (Fig. 2A). During phase 2, all three drugs produced a dose-dependent suppression of flinching response (Fig. 2B).
Intrathecal coadministration of MK801 and NBQX with a fixed dose of gabapentin in phase 1 increased the antinociceptive effect of MK801 and NBQX (Fig. 3A, 4A). Isobolographic analysis revealed a synergistic interaction after the concurrent delivery of mixtures of gabapentin-MK801 and gabapentin-NBQX during phase 2 in the formalin test. These experimental ED50 values were significantly lower than those of the theoretical ED50 values (Fig. 3B, 4B) with a total fraction
value of less than 1, indicating a synergistic interaction (Table 1).
Intrathecal gabapentin, MK801 and NBQX did not cause any change of motor tone.
DISCUSSION
In the current work, intrathecal gabapentin was active only against the phase 2 flinching response, which implicates that gabapentin may be effective in attenuating the injury-induced hyperalgesia without affecting acute nociception at the spinal level. These results were consistent with those of previous experiments (2-4). Although the antinociceptive mechanisms of gabapentin remains unclear, several targets have been proposed. It has been reported that gabapentin increases the concentration, the rate of synthesis and the release of GABA (1). But intrathecal GABAA or GABAB receptor antagonists failed to reverse the antinociception of gabapentin (5). On the other hand, NMDA and AMPA receptors were reported to be involved in gabapentin-induced antinociception (5). Additionally, gabapentin has been shown to bind specifically to the α2δ subunit of voltage-sensitive calcium channels (20).
In this study, intrathecal MK801 and NBQX resulted in a dose-dependent inhibition of flinching response in both phases. These observations suggest that NMDA receptors and AMPA receptors may be active in the modulation of the injury-induced hyperalgesia as well as acute nociception at the spinal level.
EAAs, such as glutamate and aspartate, may play an important role in nociceptive transmission in the dorsal horn of the spinal cord (6). These EAAs have been considered to facilitate spinal sensory transmission and contribute to the enhanced excitability of dorsal horn neurons through NMDA receptors and non-NMDA receptors (8, 9). The NMDA receptors exist in the substantia gelatinosa of the dorsal horn and are postsynaptic to an interneuron mediating an excitation which is responsible for the spinal nociceptive processing, including injury-induced hyperalgesia (21-23). Thus, NMDA antagonists may attenuate the noxious inputs in a tonically active state such as the phase 2 response of the formalin test. On the other hand, AMPA receptors are present in the superficial lamina of the dorsal horn and mediate excitatory transmission involving acute nociceptive inputs (24, 25). Hence, AMPA antagonists may suppress acute excitation induced by high intensity stimuli such as the phase 1 response of the formalin test. Interestingly, in this study, spinal NMDA antagonist and AMPA antagonist reduced not only the phase 1 response but also the phase 2 response. These findings suggest that the NMDA receptor and the AMPA receptor may be involved in the regulation of the injury-induced hyperalgesia as well as acute nociception in the spinal cord. The fact that NMDA antagonist blocked the release of substance-P (26) supports the phase 1 antinociception of MK801 observed in this study. Additionally, the phase 2 response seems to result from a continuous afferent input, which is produced in phase 1. Thus, as the phase 1 component of the formalin stimulus is gradually reduced by AMPA antagonist, the phase 2 response might also be decreased. This effect was observed during phase 2 in this study with AMPA antagonist.
Although NMDA antagonist and AMPA antagonist attenuated both acute nociception and injury-induced hyperalgesia, the relative effectiveness of the drugs was noted. In MK801-treated rats, ED50 in phase 2 was lower than that in phase 1. Moreover, the ED50 value of NBQX in phase 1 was similar to that in phase 2. These findings suggest that the NMDA antagonist appears to be much more effective on injury-induced hyperalgesia than on acute nociception. Additionally, AMPA antagonist shows similar effects for acute nociception and injury-induced hyperalgesia. However, the results observed in the present study are both consistent (11, 13, 15) and inconsistent with previous data (7, 10, 12, 14). This discrepancy may be caused by the different types of tested stimuli, kinds and doses of drugs administered and the relative affinity or selectivity of the drugs.
According to a fixed dose analysis performed in phase 1, intrathecal gabapentin increased the antinociceptive effect of MK801 and NBQX. Isobolographic analysis performed in phase 2 indicated that intrathecal gabapentin showed a synergistic effect with MK801 as well as with NBQX. These results illustrate that spinal gabapentin can augment the antinociceptive action of MK801 and NBQX for injury-induced hyperalgesia as well as acute nociception. Previous studies have shown that intrathecal gabapentin reinforced the effects of clonidine or neostigmine for acute nociception (18). A synergistic interaction has been reported between gabapentin and other analgesics, such as clonidine, naproxen and morphine in numerous nociceptive conditions (16, 27, 28). Several mechanisms would be possible for this synergistic interaction. First, drugs may interact by altering the kinetics of each other. One agent may alter the actions of the other agent at the receptor or channel, thereby leading to synergism. Second, functional interaction may result from distinct drug effects at separate anatomic sites that may act independently and together to inhibit spinal nociceptive processing (29). Thus, a decrease in excitatory neurotransmission with MK801 and NBQX accompanied by inhibiting calcium channels with gabapentin in the spinal cord may lead to synergism. This mechanism was observed in the synergistic interaction of gabapentin and non-NMDA antagonist in a neuropathic pain model of rats (30). Third, synergism may stem from the characteristics of action of gabapentin. Gabapentin would promote the activity of inhibitory neurons following inflammatory injuries (31) and thus the combination of gabapentin with MK801 or NBQX may result in a greater reduction of transmission of nociceptive signals, which may develop into synergism.
Altogether, intrathecal MK801 and NBQX, but not gabapentin, reduced the phase 1 flinching response of the formalin test, and all three drugs reduced the phase 2 flinching response. Intrathecal gabapentin also increased the effects of MK801 and NBQX during phase 1 of the formalin test and interacted with MK801 and NBQX in a synergistic fashion during phase 2.
These results suggest that spinal NMDA and AMPA receptors may be involved in the regulation of the injury-induced hyperalgesia as well as acute nociception. Concurrent delivery of gabapentin with either NMDA antagonist or AMPA antagonist may well have virtue in dealing with pain, as it serves to reduce the required dose of each drug





 XML Download
XML Download