

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Apo2L/TRAIL (Apo2 ligand or tumor necrosis factor (TNF)-related apoptosis-inducing ligand) was discovered by its sequence homology to TNF and CD95L (1, 2). Recombinant soluble human Apo2L/TRAIL is a candidate for clinical investigation in cancer therapy because it induces apoptosis in a broad spectrum of human cancer cell lines but not in many normal cells, and exhibits potent anti-tumor activity without normal tissue toxicity in various cancer xenograft models (3-7). To date, four homologous, but distinct, human TRAIL (hTRAIL) receptors have been identified, with two (DR4/TR1 [8] and DR5/TR2 [9-11]) having the ability to initiate the apoptosis signaling cascade after ligation, and two (TRID/DcR1/TR3 [9, 11, 12] and TRUNDD/DcR2/TR4 [13-15]) lacking this ability. Because they lack the ability to directly signal cell death, TR3 and TR4 have been hypothesized as being protective receptors, either by acting as decoy receptors (8, 14, 15) or via transduction of an antiapoptotic signal (13).
Glioblastoma multiforme (GBM) is the most malignant and common brain tumor, comprising 23% of all primary brain tumors in adults. GBM tumors are refractory to all current therapeutic approaches, including surgery, radiotherapy, and chemotherapy (16). Human glioma cells express TR2 and undergo apoptosis upon TRAIL ligation in vitro (17, 18). Local injection of TRAIL exerted strong antitumor activity on intracranial human malignant glioma xenografts in athymic mice in the absence of neurotoxicity (6). In combination with conventional DNA-damaging chemotherapy, TRAIL showed synergistic cytotoxicity for human gliomas in vivo and in vitro (19). Therefore, systemic or intracranial TRAIL treatment may be a promising approach for human GBM tumors. However, one potential drawback to these findings was that large amounts of soluble TRAIL were required to inhibit tumor formation. This may be due to the pharmacokinetic profile of soluble TRAIL that indicated that after intravenous injection the majority of the protein is cleared within 5 hr (4). Increasing the in vivo t1/2 of soluble rTRAIL or developing an alternative means of delivery may increase the relative tumoricidal activity of TRAIL such that larger, more established tumors could be eradicated as efficiently as smaller tumors. The results presented in this work describe the production of an adenoviral vector engineered to carry the gene for hTRAIL. After infection, TRAIL protein was detected, leading to the induction of apoptosis in human malignant glioblastoma cells in vitro as well as significant antitumor activity in vivo. These results demonstrate the potential therapeutic utility of adenoviral-mediated delivery of TRAIL gene.
MATERIALS AND METHODS
Production of adenovirus encoding the hTRAIL gene
The cDNA for hTRAIL was obtained from Dr. Hideo Yagita (Juntendo University, Tokyo, Japan) (20). To generate E1-deleted recombinant adenoviral vector encoding hTRAIL (Ad.hTRAIL), hTRAIL cDNA was introduced into the shuttle plasmid, pAvCvSv, under the transcriptional control of the cytomegalovirus (CMV) immediate early enhancer/promoter (21). The recombinant shuttle plasmid was co-transfected with the E1-deleted adenovirus serotype 5 genome, pJM17 (22), into transformed human embryonic kidney cells, designated 293 cells (23). A E1-deleted recombinant adenovirus, containing a reporter β-galactosidase (β-Gal) gene with CMV promoter (Ad.lacZ) was used for concurrent control. Recombinant adenoviruses were amplified on 293 cells and purified by two centrifugation steps on cesium chloride gradients (24). Viruses were dialyzed against 10 mM Tris HCl pH 8.0, 1 mM MgCl2, 10% glycerol and stored at -80℃ until use. The number of viral particles was assessed by measurement of the optical density at 260 nm.
Cell culture
The p53-wild-type human malignant glioma cell lines U87MG and the p53-mutated U373MG were obtained from the American Type Culture Collection. Each of these cell lines was maintained in growth medium consisting of RPMI1640 supplemented with 10% fetal celf serum (Life Technologies, Calsbad, CA, U.S.A.), and the following anti-microbial agents: 100 IU/mL penicillin and 100 µg/mL streptomycin. Cultures were maintained at 37℃ in a humidified atmosphere with 5% CO2 in air and subcultured every 4 to 7 days with 0.25% trypsin in HBSS (Life Technologies).
Adenoviral infection
Cells were cultured and permitted to adhere for at least 12 hr before adding adenovirus. Before infection, cells were washed with PBS, and then the vectors were added at the indicated number of multiplicity of infection (MOI)/cell in PBS. After 30 min, cells were washed with PBS and incubated in complete medium for the remainder of the assay.
Western blotting and ELISA
Cells were cultured and harvested on second and third day after infection of Ad.hTRAIL and treated with lysis buffer (0.5% sod. deoxycholate, 0.5% Triton X-100, 50 mM Tris, pH 7.4, 150 mM NaCl, 62.5 mM sucrose, 5 mM EDTA, 1 mM PMSF). Protein concentrations of the lysates were determined by the colorimetric bicinchoninic acid analysis (Pierce, Rockford, IL, U.S.A.). Equal amounts of protein were separated by SDS-PAGE, transferred to nitrocellulose membrane (Novex, San Diego, CA, U.S.A.). The membrane was incubated with the anti-PARP (BD Biosciences, San Jose, CA, U.S.A.), or anti-hTRAIL mAb (BD Biosciences). For detection of secreted TRAIL, the medium of the TRAIL-expressing cell cultures were measured by ELISA (BD Biosciences).
Nuclear morphology for apoptosis
Cells were cytocentrifuged, stained in 4 µg/mL Hoechst 33342 for 30 min at 37℃ and fixed for 10 min in 4% paraformaldehyde.
MTT assay after Ad.hTRAIL infection
Cells were infected with 10, 30, 100, 300, and 1,000 MOI of Ad.hTRAIL; 10% (volume/volume) of stock MTT solution (3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide) (Sigma, St. Louis, MO, U.S.A.) dissolved in medium at a concentration of 5 mg/mL was added to the medium directly, and cells were incubated for 30 min at 37℃, to allow cell-mediated reduction of MTT, to assay cell viability. The medium was aspirated and cells were washed with phosphate-buffered saline. DMSO (0.5 mL, equal to the volume of medium) was added to solublize the dye. Absorbance was measured at 570 nm.
Assessment of antitumor activity in a SCID mice
In vivo assessment of the effect of Ad.hTRAIL on glioma growth was performed using a SCID mice (6-8 weeks old; Daehan Laboratory Animal Center, Korea) by subcutaneous inoculation of 1×106 U87MG glioma cells into the dorsal flank of each mouse. This dose of cells produces virtually 100% tumor take in the SCID mouse system in our laboratory. To examine the apoptotic action of Ad.hTRAIL, 10 days after tumor implantation, animals were randomly chosen to receive either vehicle or 1×1010 particles of Ad.hTRAIL per mouse administered by intratumoral injection a volume of 100 µL. After 3 days the mice were sacrificed and the tumor region was excised. Histological analysis was followed. Furthermore, to determine the suppression of tumor growth of Ad.hTRAIL, animals were administered with Ad.hTRAIL or Ad.lacZ. Tumor volumes were measured every other day for 30 days.
Histological procedures and TUNEL assay
The excised tumor tissues were fixed in 10% neutral-buffered formalin and embedded in paraffin. Five µm thick sections were stained with hematoxylin-eosin for histological examination. Apoptosis induced in tumor tissues was measured using the TUNEL assay (Apoptosis Detection Kit, Chemicon, Temecula, CA, U.S.A.). Tissue sections, 5 µm thick, were deparaffinized and rehydrated through a series of graded alcohols. The sections were processed in 0.05 M sodium citrate buffer (pH 6.0) and heated in a microwave for 10 min for antigene retrieval. Samples were subjected to the reaction with terminal deoxynucleotidyl transferase in the presence of digoxigenin-conjugated nucleotide substrate at 37℃ for 30 min. After the reaction was stopped, the slides were incubated with anti-digoxigenin antibody that had been conjugated with peroxidase. The peroxidase was visualized with DAB (3,3'-diaminobenzidine tetrahydrochloride). Sections were counterstained with Mayer's hematoxylin and then coverslipped.
RESULTS
Production of TRAIL-encoding adenovirus and TRAIL gene expression
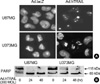
The cDNA encoding full-length hTRAIL was inserted into the E1 region of a replication-deficient Ad5 construct under the control of the CMV immediate early promoter. The engineered TRAIL-expressing adenovirus was infected into U87MG and U373MG cells, and the cellular proteins were separated by SDS-PAGE to assay for TRAIL expression by Western blotting. Amino acid sequence analysis of the TRAIL cDNA predicts a weight of 32.5 kDa for TRAIL monomers (1, 2). Treatment of cells with Ad.hTRAIL resulted in a strong TRAIL-specific band according to particle number of infection and period for incubation (Fig. 1). To confirm the secretion of TRAIL in the medium of the TRAIL-expressing U87MG cell cultures, ELISA showed that the concentration of TRAIL is 235±28 pg/mL.
TRAIL-induced apoptosis in human malignant glioma cells
To test whether treatment with the Ad.hTRAIL, which is known to elicit apoptosis in a variety of transformed or malignant cells (3, 4), would similarly elicit apoptosis in cultured cancer cells, the cell-killing effect of the Ad.hTRAIL was analyzed by quantifying apoptotic cells by measuring cell viability via MTT assay (Fig. 2). The results show a significant difference in cell killing between lines in response to treatment with TRAIL-expressing vectors versus control vectors. Although MTT assay of the tumor cells infected with Ad.hTRAIL as presented in Fig. 2 indicates the amount of cell death, it does not discriminate between apoptotic and necrotic cell death. Previous reports have demonstrated that TRAIL-induced cell death occurs through an apoptotic mechanism characterized by the activation of a cascade of intracellular proteases and the cleavage of numerous intracellular proteins (25-27). To confirm that the tumor cell death following Ad.hTRAIL infection was mediated through an apoptotic mechanism, nuclear fragmentation and cellular protein cleavage were examined. Morphological studies using Hoechst nuclear staining revealed nuclear condensation in cells treated with Ad.hTRAIL (Fig. 3A). U87MG and U373MG cells were incubated with Ad.hTRAIL for 30 min, cell lysates were prepared at various times after infection, and the cellular proteins were separated by SDS-PAGE for Western blot analysis of PARP cleavage as substrate for caspase-3 (Fig. 3B). These results demonstrated that treatment with the TRAIL gene effectively elicited apoptosis in cultured malignant glioma cells.
Tumor growth suppression by intratumoral delivery of the TRAIL gene in vivo
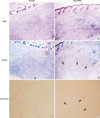
To further test the antitumor effect of the TRAIL gene, human malignant glioma xenografts were established in SCID mice by inoculating U87MG cells subcutaneously into the dorsal flanks of mice as described above in "Materials and Methods." Histochemical analysis by H&E 3 days after intratumoral injection of Ad.hTRAIL into preexisting tumor revealed extensive disappearance of tumor cells in center which demonstrated that intralesional administration of the TRAIL gene effectively killed tumor cells (Fig. 4B). Ad.lacZ-infected tissue showed well preserved and proliferated tumor cells (Fig. 4A). Furthermore, TUNEL assay demonstrated that the Ad.hTRAIL-infected cells showed positive stain for apoptotic cells at periphery (Fig. 4D). while Ad.lacZ-treated cells were not stained (Fig. 4C). To demonstrate whether Ad.hTRAIL was able to suppress growth of preexisting tumor, we gave a single intratumoral injection of Ad.hTRAIL or Ad.lacZ at 7 days after tumor implantation. Ad.hTRAIL significantly suppressed tumor growth (p<0.01) (Fig. 5). The growth of the tumors treated with Ad.hTRAIL was well suppressed until day 30 on average.
DISCUSSION
Human malignant glioma cells are highly resistant to multiple pro-apoptotic stimuli, including irradiation and cytotoxic drugs. Although loss of p53 function and alterations in cell cycle control genes have been attributed a role in this radiochemoresistant phenotype, the precise molecular pathways mediating glioma cell resistance to radiotherapy or chemotherapy have not been elucidated. Therefore, death ligand, TRAIL is potentially valuable agent for the induction of glioma cell death (17, 28). TRAIL has become an attractive molecule for the treatment of cancers because it specifically kills tumor cells (3, 4). Recently it is reported that TRAIL induced apoptosis through death receptor 5 (DR5) and was mediated by caspase-8-initiated extrinsic and intrinsic mitochondrial pathways in sensitive glioma cell lines. Furthermore, TRAIL alone or in combination with chemotherapeutic agents, induced apoptosis in primary tumor cultures from patients with malignant gliomas, reinforcing the potential of TRAIL as an effective therapeutic agent for malignant gliomas (33). The tumor-specific activity of TRAIL was extended in vivo with the observation that treatment of SCID and nude mice bearing human tumors with soluble TRAIL significantly inhibited tumor outgrowth without any observable toxic side effects to the host (5-7). This inhibition of tumor outgrowth, though, required high amounts of recombinant TRAIL given over several days shortly after tumor implantation. Pharmacokinetic analysis revealed that soluble TRAIL given to mice intravenously displayed an elimination t1/2 of just under 5 hr (4). An alternative approach would be to administer TRAIL locally, where it would exist at a greater concentration and have a better chance of significantly inducing tumor cell death. Such localized, intratumoral injections of soluble TRAIL would, however, be limited in that only a relatively small volume could be administered, suggesting that a potentially suboptimal amount of TRAIL protein would be used. In contrast, Ad.hTRAIL can be produced at high titers, such that small volumes would contain high numbers of infectious adenoviral particles carrying the hTRAIL gene.
In the study reported here, we asked whether introducing the TRAIL gene via adenoviral vector directly into malignant glioma cells would result in the expression of biologically active molecules that could effectively kill malignant cells in vitro and in vivo. The data presented in this study describe the generation of an adenoviral vector engineered to carry the gene for hTRAIL. Ad.hTRAIL infection resulted in the transcription and translation of the transferred hTRAIL gene into functional TRAIL protein that, when expressed on the cell surface, induced apoptotic death in malignant glioma cells. One report described that adenoviral TRAIL gene transfer is ineffective treatment strategy for malignant glioma (29). In this study, however, adenoviral TRAIL gene transfer showed apoptosis in human glioma cells and even induced cell death in TRAIL-resistant glioma cells (U373MG). We have found that treatment with TRAIL-expressing vectors produced a substantial amount of TRAIL on the surface of the cells and secreted soluble TRAIL in the medium of the TRAIL-expressing cell cultures. These results suggest that the apoptotic activity of TRAIL produced by adenoviral vector is elicited via both of membrane-bound TRAIL and soluble TRAIL. The effects of soluble TRAIL may be dose-dependent, conformation-dependent, or both. It has been reported that trimerized recombinant TRAIL is more effective than the monomer form; thus, conformational differences in soluble TRAIL proteins may also dramatically affect their activities (30). Our data suggest that transmembrane expression is indeed a highly potent strategy for expressing TRAIL. Why transmembrane expression of TRAIL is so highly efficient for inducing target cell apoptosis is unclear, although possibilities include transmembrane TRAIL undergoing oligomerization at the cell membrane following binding to its receptor on target cells.
The present study examined the possible therapeutic value of an adenovirus encoding TRAIL gene for the treatment of malignant glioma. This virus has previously been shown to kill melanoma, breast carcinoma cell lines (27), prostate and bladder carcinoma cells (31), and lung, colon cancer cell lines (32). In this study, we demonstrated that Ad.hTRAIL induces cell death of malignant glioma. Our data confirm recent studies demonstrating antitumor activity of Ad.hTRAIL, and suggest TRAIL gene transfer as a potential alternative strategy for the efficient delivery of TRAIL.



 XML Download
XML Download