

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
One of the major mechanisms for controlling blood circulation is the regulation of vascular resistance through the change in vascular tone. Vascular tone is affected by the changes in extracellular and intracellular pH (pHo and pHi) and it influences blood flow. It is well known that vascular response during acidosis causes significant alteration in blood circulation. Tian et al. (1) reported that hypercapnic acidosis induced vasodilation. And it has been known about role of pHo in vascular function; extracellular acidosis ([acidosis]o) induces relaxation and extracellular alkalosis ([alkalosis]o) develops vice versa. Since cerebral arterial relaxation by [acidosis]o was maintained when the level of pHi was kept constant (1-3), pHo is also very important for the regulation of vascular function.
pH has been shown to modulate calcium influx and to regulate intracellular calcium concentration (4-6). As known well, the change in the intracellular Ca2+ concentration ([Ca2+]i) is responsible for smooth muscle contraction, and H+ also is an important regulator of contractility. In addition, both ions are connected to and changed by each other. Physiologically the alteration of these ions, due to hypoxia or other stimuli, can also affect the distribution of blood and the maintenance of blood pressure. Therefore, the regulation of [Ca2+]i by pH is very important in vascular smooth muscle. In smooth muscle, changes in [Ca2+]i are critical factors for contraction or relaxation and the initiation of other cellular responses. Changes in [Ca2+]i reflect an influx of Ca2+ via the sarcolemmal channels and/or a release from cytosolic Ca2+ stores (sarcoplasmic reticulum, SR) through the processes of Ca2+-induced Ca2+ release (CICR) or through the inositol 1,4,5-trisphosphate (InsP3)-mediated Ca2+ release (7). Among them, Ca2+ influx pathway through voltage-dependent Ca2+ channel (VDCC) in arterial smooth muscle is important for the contractile response, including the maintenance of the basal tone (8, 9). In basilar arteries of rabbit, two types of Ca2+ channels (T and L-type) were reported (12-16). Among them, L-type Ca2+ channel largely contributes to the Ca2+ influx in vascular smooth muscle. Although we could not find T-type Ca2+ channel or test the effects of pHo on that channel in this study, L-type Ca2+ channel is also known to be more sensitively affected by H+ ions in cerebrovascular artery than T-type Ca2+ channel (16, 17, 23). Conclusively, Ca2+ channel especially L-type Ca2+ channel regulations by pH could clearly account for the changes in [Ca2+]i and also in tone of vascular smooth muscle (10-12). In vascular smooth muscle, the regulation of VDCC by pH was reported in pial and porcine coronary arteries (4, 5), including basilar arteries of guinea pigs (16).
Although the effects of pHo on Ca2+ channel of vascular smooth muscle were reported in several cases, the more study about the regulation of Ca2+ channel and contraction by pHo in rabbits basilar arteries is still needed. In addition, some reports suggested that the regulatory effect of Ca2+ channels by pHo was observed, but there was no significant effect on contractile responses (4, 5, 18). For these reasons, this study was designed to verify the role of pHo in the relation between the contractile response and Ca2+ channel in basilar arteries of rabbits.
MATERIALS AND METHODS
Cell Isolation
Single smooth muscle cells were enzymatically isolated from the rabbit basilar arteries. Rabbits (white rabbits from NewZealand, 1.5-2 kg) were anesthetized with sodium pentobarbital (40 mg/kg, i.v.) and exsanguinated. The brain was rapidly removed and placed in phosphate-buffered cold Tyrode's solution containing (in mM): NaCl 145, KCl 5, MgCl2 2, CaCl2 2, glucose 10, NaH2PO4 0.42, Na2HPO4 1.81, HEPES 10, pH 7.4. Then the segment of basilar artery was gently dissected from the surface of the brain stem and was placed in a Ca2+-free phosphate-buffered Tyrode's solution (CaCl2 was omitted from the above compositions). Tunica adventitia, surrounding connective tissue, and side branches were removed gently under a stereomicroscope. The artery was longitudinally dissected and rinsed to remove the residual blood in vessel. The artery was first moved to a nominally Ca2+-free Tyrode's solution containing (in mM): NaCl 145, KCl 5, MgCl2 2, glucose 10, HEPES 10 (pH 7.4 with Tris). It was then cut into small pieces and the segments were stored at 4℃ for 15 min. Collagenase (1.5-1.8 mg; Wako pure chemicals) and dithioerythreitol (0.5 mg; Sigma Chem. Co., St. Louis, MO, U.S.A.), bovine serum albumin (2 mg; Sigma), and trypsin inhibitor (1 mg; Sigma) were dissolved into 1 mL of Ca2+-free Tyrode's solution. Then the arterial segments were incubated in this enzyme cocktail at 35℃ for 20-25 min. After collagenase treatment, segments were transferred to modified K-B solution (see the composition in solution and drug subsection below) and single cells were dispersed by gentle agitation with glass pipette. Isolated single cells were stored at 4℃ until use.
Electrophysiological Recording
An aliquot of single basilar arterial smooth muscle cells in suspension was added to the recording chamber (0.1 mL) mounted on an inverted microscope (Olympus, IMT-2, Japan). Solutions were superfused through the chamber by gravity at the rate of 2-3 mL/min. Experiments were performed at room temperature. Single cells were voltage-clamped, and membrane currents were measured using the conventional and perforated whole-cell configurations of patch-clamp technique with a patch-clamp amplifier (Axon Instruments, Axopatch-1D, U.S.A.; 19, 20). Patch pipettes were made from borosilicate glass capllaries (inner diameter: 1.5 mm), pulled on a two-step vertical puller (Narishige, PP-83, Japan), and fire-polished with a microforge (Narishige, MF-83); the pipettes had resistances of 3-5 MΩ. To measure whole-cell currents after gigaseal formation, access to the cell interior was obtained by rupturing the membrane at the tip of the pipette with additional negative pressure. Series resistance was not compensated.
Membrane currents were monitored on oscilloscope (Philips, PM 3350, Japan), and data were digitized on-line with an analog-to-digital interface (Axon Instruments, Labmaster TL-1 DMA interface, U.S.A.). Data that were digitized on-line were stored in an IBM-compatible computer. Whole-cell currents were filtered at 5.0 kHz and digitized. All data were analyzed with pClamp 5.5.1 software (Axon Instruments). Leakage current subtraction was performed on data, if necessary.
For the perforated mode patch clamp, amphotericin was dissolved in dimethyl sulfoxide (DMSO) as a stock solution (0.15 mg/2 mL) and added to the back-filling pipette solution (0.15 mg/mL).
Measurement of Mechanical Activity
Isolated basilar artery was placed in the HEPES-buffered Tyrode's solution, and the vessel ring was prepared (1.5 mm in width). The endothelium of the artery was not removed. Each ring was mounted vertically in organ bath containing 0.5 mL of Ca2+ physiological salt solution (PSS) containing (in mM): NaCl 135, KCl 5, CaCl2 1.8, MgCl2 1, glucose 10, and HEPES (N-[2-hydroxyethyl] piperazine-N'-[2-ethanesulphonic acid]) 10 which was adjusted to pH 7.4 with NaOH. For the 50 mM of K+ Ca2+ PSS solution, equimolar concentration of NaCl was replaced by KCl. The Ca2+ PSS solution was maintained at 37℃ and was continuously aerated with 100% O2. The isometric tension was measured with a force-transducer (Harvard, U.S.A.). Each ring was stretched passively to a resting tension of 300 mg. After equilibration for more than 90 min, contractile response of the strip to the solution containing 50 mM K+ was repeated two or three times until the responses were reproducible.
For the studies of mechanical contractions under various levels of pH, each solution was titrated at 37℃ to various levels of pHs. Then these solutions were pre-incubated with continuous aeration by 100% O2 in water bath (37℃) until the application to the myograph.
Solutions and Drugs
Ca2+ PSS containing (in mM) NaCl 135, KCl 5, CaCl2 1.8, MgCl2 1, glucose 10, and HEPES 10, was adjusted to pH 7.4 with NaOH. Modified KB solution (21) containing (in mM) L-glutamate 50, KCl 50, taurine 20, KH2PO4 20, MgCl2 3, glucose 10, HEPES 10, and ethyleneglycol bis-(β-aminoethyl ether-N,N,N',N'-tetraacetic acid (EGTA) 0.5, was adjusted to pH 7.4 with KOH. For conventional whole-cell recording, the following intracellular solution containing (in mM): CsCl 110, TEA 20, EGTA 10, HEPES 10, Na2ATP 3, and MgCl2 3.5, was adjusted to pH 7.3 with TRIZMA or CsOH. For perforated patch experiments, electrodes were filled with solution containing (in mM): CsCl 110, TEA-Cl 20, EGTA 10 and HEPES 10 was adjusted to pH 7.3 with TRIZMA or CsOH. All drugs used in this study were purchased from Sigma.
RESULTS
Effects of External pH (pHo) on High K+ (50 mM)-induced Contraction in Rabbit Basilar Arteries
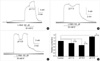
To exclude the effects of nitric oxide (NO) released from endothelial cells, nitro-L-arginine (L-NNA), an inhibitor of NO synthase, was used. In all experiments, 100 µM L-NNA was pretreated before the application of high K+ solution or the agonist. The pretreatment with L-NNA increased basal tone slightly by 1.1±0.2 mN (n=6, p>0.05, Fig. 1A). When high K+ solution was applied at the normal pHo, the arterial tone was greatly enhanced by 11.4±0.6 mN (n=19) and maintained at a sustained level (Fig. 1, 2). These tonic contractions was almost completely blocked by nicardipine (1 µM) to 7±3.9% (n=7) of the control (Fig. 5A). To elucidate the effects of pHo on K+-induced contraction, pHo was changed from 7.4 to 7.9 [alkalosis]o or to 6.8 [acidosis]o. When the pH of the bath solution was changed from 7.4 to 7.9, the high K+-induced contraction was enhanced reversibly to 128±2.1% (n=13, p<0.01, Fig. 1A, D). However, K+-induced contraction decreased to 87±1.0% and to 73±1.3% of the control at pHo 7.0 and 6.8 in a reversible manner, respectively (n=4, Fig. 1B-D). The effects of [alkalosis]o or [acidosis]o on the K+-induced contraction were also observed in the same tissue. As shown in Fig. 2A, B, enhancing and suppressing effects of pHo on high K+-induced tonic contractions were observed in the same tissue. The enhancing effects of [alkalosis]o on high K+-induced tonic contraction were also observed under the pre-application of pHo 7.9 (Fig. 2C).
Effects of pHo on histamine-induced Contraction in Rabbit Basilar Arteries
As shown in Fig. 3A, histamine (1-20 µM) produced a contraction in a concentration-dependent manner, and maximal contraction was observed approximately at 10 µM of histamine (22). In a few cases, oscillatory small phasic contractions were observed after the application of histamine (Fig. 3B). Histamine (10 µM) developed a tonic contraction with the amplitude of 11±0.6 mN (n=17, Fig. 4), which was blocked by post-application of nicardipine (1 µM), known as a blocker of Ca2+ channel (L-type) (Fig. 3C). From the 14 tested tissues, histamine-induced sustained contractions were suppressed by nicardipine (1 µM) to 21±7.3% of the control (Fig. 5B). These results suggest that Ca2+ influx through Ca2+ channel (L-type) might be important in the histamine-induced tonic contraction. Histamine-induced contractions were enhanced or suppressed under the condition of [alkalosis]o or [acidosis]o to 134±5.7% and to 27±7.6% of the control in a reversible manner (Fig. 4; n=5, 6, p<0.01), respectively.
Effects of pHo in the presence of nicardipine on High K+- and histamine-induced Contraction in Rabbit Basilar Arteries
Since high K+- and histamine-induced sustained contractions were affected by nicardipine and pH, we also studied about the possibility whether the regulatory effects of pHo on high K+- and histamine-induced contractions are associated with Ca2+ channel regulation (Fig. 5). As shown in Fig. 5A, high K+-induced contraction is absolutely nicardipine-sensitive and it is not significantly reversed by [alkalosis]o (9±5.8% of the control, n=7, p>0.05). And also nicardipine-sensitive portion of histamine-induced contraction was not significantly reversed by [alkalosis]o in Fig. 5B (28±10.2% of the control, n=13, p>0.05). These results might imply apparent involvement of Ca2+ channel regulation by pH in high K+- and histamine-induced sustained contractions. Therefore, we studied Ca2+ channel regulation by pH using basilar arterial single myocytes in next step.
Effects of pHo on Ca2+ Channel Current (IBa) in Basilar Arterial Myocytes of Rabbits
Ca2+ current (ICa) was recorded at the physiological calcium concentration of 2 mM under conventional whole-cell configuration (Fig. 6A, B). From a holding potential of -80 mV, depolarization above -30 mV produced an inward current showing a peak current at 0-10 mV, and a reversal around 50 mV (Fig. 6B, open circle). The effects of 1 µM nicardipine on ICa were shown in Fig. 6A, B. In current/voltage (I/V) relation, the peak current of ICa was markedly suppressed at the whole test potential range after the application of nicardipine (1 µM) (Fig. 6B; 6.9±8.4% of the control at 10 mV, n=4). Since the calcium channel current recorded in replacing 2 mM Ca2+ with 10 mM Ba2+ (IBa) is much larger in amplitude, direct effect of pHo on IBa was studied under perforated-patch clamp configuration. From our unpublished data, currents recorded under perforated mode are not different from those under conventional whole-cell mode. The membrane potential was held at -80 mV, and step 0 mV depolarizing pulse was applied for 500 msec every 15 sec. When the peak amplitude of IBa reached a steady-state level, the effects of [alkalosis]o and [acidosis]o on IBa were observed. As shown in Fig. 6C, D, the peak amplitude of IBa increased or decreased under the condition of [alkalosis]o or [acidosis]o. When [alkalosis]o was developed in bath solution, the peak current increased by 35±2.1% (n=3); however, [acidosis]o decreased the peak current by 41±8.8% at 0 mV (n=3).
DISCUSSION
The results obtained from this experiment suggested that enhancing or suppressing effects of pHo change on the vascular tone are associated with the modulation of VDCC in rabbit basilar arteries. To elucidate the involvement of Ca2+ channel activity in the regulation of high K+- and histamine-induced tonic contraction by pHo, we studied the effects of [alkalosis]o on those contractions in the presence of nicardipine (1 µM). As shown in Fig. 5A, B, nicardipine-sensitive tonic contractions of high K+ and histamine was not significantly recovered by [alkalosis]o. That strongly means that VDCCL is a major target for the regulation of high K+-induced tonic contraction by [alkalosis]o. This experimental procedure enabled us to find the possible mechanism of Ca2+ channel modulation of those contractions by pH.
As a potent modulator of cerebrovascular tone, histamine causes a cerebral vasoconstriction and dilation (8, 9). To date, histamine in vascular smooth muscles has been known to provoke following representative changes in the cytosolic level. Firstly, it can induce IP3 production in smooth muscle, thereby resulting in IP3-induced mobilization of Ca2+ from SR (8). Secondly, it depolarizes vascular smooth muscle cells through the activation of nonselective cation channels (NSCC), thereby increasing Ca2+ influx through VDCC resulting in tonic contraction (8, 9, 24, 29, 30). Thirdly, it can also directly increase Ca2+ current in some smooth muscles (31, 32). Among them, regulation of Ca2+ current is very important since Ca2+ current is associated with tonic contraction. These tonic contraction is simultaneously linked to depolarization and a primary physiological role of agonist-induced depolarization is an activation of VDCC with subsequent influx of Ca2+ and then stimulation of contraction (24-27). In cerebral artery, it was reported that histamine and high K+ produced a sustained tonic contraction that was blocked by 1 µM of nicardipine. Gokina and Bevan (8, 9) reported that Ca2+ influx through VDCC might be responsible for over the 80% of histamine-induced sustained contraction (26). The same result was also reported in human cerebral arteries (33) and observed in this study (Fig. 3C, 4, 5B). These results suggests that the maintenance of histamine-induced sustained tonic contraction might also be responsible for the activation of Ca2+ channel (L-type) through depolarization. Meanwhile, high K+ is also well known to produce depolarization and associated dihydropyridine-sensitive tonic contractions (Fig. 5A) (8). These observations suggest the possibility that the regulation of those tonic contraction through modulation of Ca2+ channel.
Although we suggested the functional role of Ca2+ channel regulation by pHo, some differences in the extent of the effects of pHo between pHo effect on the actions on the Ca2+ channel (L-type) current and that on the K+-induced contraction were also observed. [acidosis]o decreased the peak current of IBa by 41% of the control but reduced K+-induced contraction by 27% of the control (Fig. 1D, 6D). Meanwhile, [alkalosis]o increased the peak amplitude of IBa by 35% of the control, but enhanced K+-induced contraction to 23% of the control (Fig. 1D, 6C). A greater difference was observed in the effects of [acidosis]o on histamine-induced contraction. When [acidosis]o was applied to the histamine-induced contraction, histamine-induced contraction was reduced to 27% of the control (Fig. 3). Eventually, these differences might be originated from the different extent of the activation of VDCC between the tissue and a single cell. According to the other report, high K+ solution (35-66 mM) produced membrane depolarization around -30~-20 mV (8). Under the 50 mM of high K+ solution for the K+-induced contraction, membrane potential in rabbit basilar arteries is expected to be depolarized between -30~-20 mV. Ca2+ channel (L-type) can be activated at these membrane potentials but very small portion of current (approximately 15% of the peak current at 0 mV) will be activated in I/V relation (Fig. 6B). In addition, there might be another possibility that the changes of pHo can affect different ionic conductances such as K+ channels and then lessens the effects of the pHo on Ca2+ channel (L-type). In 1998, the effects of acidosis on Ca2+-activated K+ channel (KCa channel) and ATP-sensitive K+ channel (KATP channel) in coronary artery were reported (7, 34). Although the data were not shown, we observed the effects of TEA and glibenclamide, which are known to be the blockers of KCa channel and KATP channel. Glibenclamide (10 µM) did not show any significant effects on acidosis-induced relaxation of histamine-induced contractions (n=2). However, TEA partially reversed acidosis-induced relaxation of histamine-induced contraction. Therefore, co-involvement of KCa channel activation is suggested with the modulation of Ca2+ channels by [acidosis]o in histamine-induced contraction. Although above several possible ionic conductances which might be involved in the regulation of histamine-induced contraction by the change in pHo were discussed, direct modulatory effect of pHo on histamine-induced contraction should also be considered. To date, in fact, most regulatory effects of pH in the regulation of vascular tone have been studied to determine the interaction between pH, [Ca2+]i and ionic conductances. From these reasons, the interpretation of the effect of pH on vascular tone should be careful until direct effect of pH is established.
As shown in Fig. 3A, histamine produced tonic contractions in a concentration-dependent manner. When various concentrations (1-20 µM) of histamine were applied to bath solution, significant contractions was recorded from 0.5 µM of histamine (Fig. 3B) and maximal contraction was observed at 10 µM of histamine. Histamine (0.5, 1, 3 µM) produced 3±0.9, 42±6.9, and 64±6.4% of the maximal contraction, respectively (n=3, data not shown). Concentration-response relation of histamine in rabbit basilar artery was already reported and our observation is in good agreement to published data (22). In Fig. 3B, some oscillatory vasomotions induced by histamine were observed in rabbit basilar arteries. To date, the physiological significance of active vasomotion in large arteries is not yet clear. Most spontaneous contraction is observed in veins but is rare in large arteries, and it may be induced by neural transmitters including hormonal vasoactive substances (37). However, oscillatory contractions are associated with oscillatory change in [Ca2+]i in vessels (35, 36). In addition, agonist-induced intracellular Ca2+ oscillations were already reported in vascular smooth muscle (37). Such intracellular Ca2+ oscillations may be responsible for the histamine-induced oscillatory vasomotion. We observed the histamine-induced oscillatory contractions in a few cases, and it is known that arteries often produce oscillatory contraction in pathophysiological conditions. Therefore, further study on the oscillatory vasomotion is needed to verify the underlying mechanism of this phenomenon.
In the present study, we tried to elucidate the involvement of VDCC in the changes in pH-induced contraction of vascular smooth muscle. However, the contribution of vascular endothelium to the pH effects on intact vessels should be taken into consideration (38). Two kinds of endothelium-derived mediators have been proposed to account for endothelium-dependent relaxation; one is nitric oxide, and the other an endothelium-derived hyperpolarizing factor (EDHF) (39-41). For these reasons, L-NNA was used to block possible additional involvement of NO-induced inhibitory influences in pHo effects on high K+- and histamine-induced contractions. However, we did not exclude the possible involvement of EDHF in the pHo regulation of contractions in this study. In 1992, Nagao and Vanhoutte suggested that NO-independent relaxation was abolished by high K+ solutions in porcine coronary artery (42). As shown in Fig. 1, [acidosis]o significantly decreased high K+-induced contractions. This findings suggest that inhibitory effects on contractions by [acidosis]o might be EDHF independent relaxation in rabbit basilar arteries. Further studies about the involvement of EDHF in inhibitory effect of [acidosis]o on histamine-induced contraction are needed. To date, regarding the modulation of pHo, most studies on the effects of pH have been done by changing the concentration of the NaHCO3 or PCO2 levels. However, in this study, HEPES-buffered PSS solution was used for recording contractile responses. Since this experiment was designed to compare the effects of pHo both Ca2+ channel current and contraction, the same HEPES-buffered PSS solution (PSS-buffered solution) was used for the studies on contraction and IBa (18).
From these results, it could be concluded that the regulatory effects of pHo on high K+- and histamine-induced tonic contractions might be partly associated with the modulation of L-type Ca2+ channel current in rabbit basilar arteries.



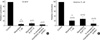

 XML Download
XML Download