

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Despite recent advances in heart failure (HF) treatment, the incidence, mortality, and economic burden associated with HF continue to grow due to an aging population and increased prevalence of risk factors including ischemic heart disease, diabetes, and obesity. Novel preventive and reparative therapies are needed. Drug development for HF has proceeded smoothly at early in vitro stages, but in many cases has failed at late stages either because the target did not produce the expected effect in whole animals or because of unexpected side effects.1) Traditional mammalian models of HF are both expensive and difficult to work with, due to the long reproduction cycle, high cost, and large space. Therefore, there is growing demand for new, simple, rapid, cost-effective ways to evaluate compounds for their effect on HF, particularly during early developmental stages.2)
The etiology of HF is diverse. In the clinical realm, chronic hypertension and ischemic heart disease are major contributing factors.3) Dilated cardiomyopathy is the single most prevalent etiology of non-ischemic HF.4) Dilated cardiomyopathy is clinically characterized by ventricular dilation and systolic dysfunction.5) As a result, there is biventricular and biatrial enlargement, elevation of left- and right-sided filling pressures, and an increase in organ and chamber weight with myocyte hypertrophy.6) Several animal models of dilated cardiomyopathy have been developed using numerous small and large animals including dog, pig, rabbit, and mouse for drug development and mechanism study.7)8)9)10) Coronary artery ligation, transgenic overexpression/knockout, pacing-induced tachycardia, pressure-overload, or cardiotoxic drugs are used to produce dilated cardiomyopathy. Doxorubicin11) or isoproterenol12) are frequently used to induce HF.
In this study, we developed a zebrafish-dilated cardiomyopathy model using short-term terfenadine treatment. Zebrafish have a similar cardiac cycle and cardiac morphology to humans, with substantial genetic conservation.13) Furthermore, zebrafish embryos are transparent, allowing easy investigation into heart structure, function, and blood flow after 48 hours post fertilization (hpf) by light microscopy.14) Zebrafish are suitable for extensive drug screening because of relatively low maintenance cost, accessibility of in vivo imaging, and tiny size.15) With these benefits, several zebrafish disease models have been introduced.16) We used terfenadine to induce non-ischemic HF. Terfenadine is a non-sedating histamine-1 receptor blocker, overdoses of which are known to induce QT prolongation and life-threatening torsade de pointes cardiac arrhythmia.17) Terfenadine increases spontaneous sarcoplasmic reticulum calcium release in cardiac myocyte.18) Calcium ions are accumulated in the cardiomyocytes and induce apoptosis.19) With this mechanism, we used terfenadine to induce dilated cardiomyopathy in zebrafish.
METHODS
Zebrafish
Zebrafish (Danio rerio) were normally maintained under a 14 hours light/10 hours dark cycle in an automatic circulating tank system and fed brine shrimp 3 times a day. Three and 4 pairs of zebrafish were set up for mating. On average, 200 to 300 embryos were generated. Embryos were maintained at 28°C in egg water as described in the zebrafish reference book. Experiments were performed with 3 days post fertilization (dpf) of hatched zebrafish embryos per experiment. Transgenic cmlc-2:GFP zebrafish, which express enhanced green fluorescent protein (GFP) in cardiomyocytes, were used to visualize myocardial cells located around both the heart chambers and the atrioventricular canal.20) The animal study was performed in accordance with international rules for animal experiments and the internationally accepted ethical principles for laboratory animal use and care. Protocols were approved by the Institutional Animal Care and Use Committee of Seoul National University (accession number SNU-150330-3).
Chemical treatment
Zebrafish at 3 dpf were transferred to 12-well plates, 10 zebrafish per well with 1 mL of phenylthiourea (PTU) solution for a treatment period in order to reduce zebrafish eye size. Depending on the experimental design, PTU solution was replaced with the same volume of PTU solution containing terfenadine or 0.1% dimethyl sulfoxide (DMSO) as the vehicle control.
Cardiac morphology
After treatment, zebrafish were mounted in 1% low melting agarose. Images were obtained using a stereomicroscope (Leica M165 FC; Leica Microsystems, Milton Keynes, UK) for morphology observation. Video imaging of the beating heart was captured by fluorescence microscope (Leica AF2000; Leica Microsystems) linked to a PC using the Leica Application Suite for Advanced Fluorescence (LAS AF) program. Ventricular size was determined by measurement across endocardial borders at end diastole. Fractional shortening was calculated using ventricular size at both end systole and diastole state. Calculating formula: where VIDd is diastolic ventricular internal dimension, and VIDs is systolic ventricular internal dimension.
Heart rate and arrhythmia rate
Atrial and ventricular rates were counted for 30 seconds with a stopwatch and a counter in cmlc-2:GFP TG zebrafish. The number of heart beats per unit of time was typically expressed as beats per minute (bpm). To analyze cardiac rhythm, atrial beats were counted for 30 seconds. Arrhythmia rate was calculated using the formula:
RNA isolation and quantitative real-time polymerase chain reaction (qRT-PCR)
For qRT-PCR analysis, total RNA was extracted from 20 larvae per treatment group using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's instructions. Isolated 1 µg of total RNA was reverse-transcribed into cDNA using M-MLV reverse transcriptase (Enzynomics, Daejeon, Korea). Quantitative PCR was performed using SYBR green qPCR Master Mix (Applied Biosystems, Foster City, CA, USA) and an ABI real-time PCR 7500 machine (Applied Biosystems) using following primers. All samples were normalized to β-actin mRNA level. Primer sequences are listed in Table 1.

Hematoxylin and eosin (H&E) staining
Zebrafish were collected, fixed in 10% neutral buffered formalin at room temperature (RT) overnight, and subjected to paraffin embedding and sectioning. Heart sections were stained with H&E.
Acridine orange staining
After 1 hour of exposure to terfenadine, zebrafish were washed twice in PTU solution and then transferred into 1 mg/mL of acridine orange dissolved in PTU solution for 10 minutes at RT.21) Larvae were then washed with PTU solution 3 times for 5 minutes each. Before examination, embryos were anesthetized with 0.016 M tricain for 3 minutes. Apoptotic cells were identified with a florescence microscope. The apoptotic cells appeared as obvious bright spots.
RESULTS
Transient terfenadine treatment reduced heart rate, inducing blood stagnation
We used terfenadine to induce QT prolongation and cardiac arrhythmia in zebrafish.1)22) Zebrafish have a single-loop circulatory system with a 2-chamber consists of an atrium receiving blood and a ventricle pumping blood to the body (Figure 1). We treated the zebrafish with 5 to 10 µM of terfenadine for 24 hours and then changed the egg water. Control zebrafish in 0.1% DMSO showed normal morphology (Figure 2A). However, terfenadine-treated zebrafish showed enlarged heart size (black arrow) and venous congestion (hollow arrow) (Figure 2B and C). Heart rate was markedly reduced even after terfenadine withdrawal (Figure 2D).
Figure 1
Morphology of 5 dpf zebrafish larvae after H&E staining.
dpf = days post fertilization; H&E = hematoxylin and eosin.
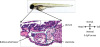
Figure 2
Transient terfenadine treatment reduced heart rate, inducing blood stagnation. (A) Representative image of control zebrafish larvae treated with 0.001% DMSO for 24 hours. (B, C) Representative images of zebrafish larvae treated with 5 µM (B) and 10 µM (C) terfenadine for 24 hours. Terfenadine-treated zebrafish showed enlarged heart size (black arrow) and venous congestion (hollow arrow). (D) Heart rates in control and terfenadine-treated zebrafish larvae (n=20 zebrafish/group).
DMSO = dimethyl sulfoxide.
*p<0.050.
Atrioventricular dyssynchrony was induced after terfenadine treatment
We used transgenic cmlc-2:GFP zebrafish, which express enhanced GFP in cardiomyocytes, in order to better visualize myocardial cells located around both the heart chambers and the atrioventricular canal. Therefore, atrium and ventricle were clearly visualized, and their contraction timing was discriminated (Figure 3A). Serial images were produced using in vivo video recording (Figure 3B). Outlines of the atrium are shown with white dashed lines, and outlines of the ventricle with red lines. The interval between images in the montage is 0.196 seconds. Compared with the controls, terfenadine-treated zebrafish showed uncoupled atrial-ventricular contraction. In control zebrafish, when the atrium was contracted, the ventricle was dilated at a well matched 1:1 ratio. In contrast, atrial contraction was not matched with ventricular dilation in the treated fish. When we measured atrioventricular ratio in the control, the ratio was approximately 1 (1:1 match). However, the atrioventricular ratio in the terfenadine-treated zebrafish was around 0.7, indicating a substantial blockage of atrioventricular conduction (Figure 3C). Atrial area was markedly enlarged by terfenadine (Figure 3D).
Figure 3
Atrioventricular dyssynchrony was induced after terfenadine treatment. (A) Representative image of cmlc-2:GFP transgenic zebrafish larvae. (B) Images taken from in vivo video recording at 4 dpf in the control (0.001% DMSO) and 24-hr terfenadine (20 μM)-treated zebrafish larvae. The interval between images in the montage is 0.196 seconds. Outlines of the atrium are shown with white dashed lines, and outlines of ventricles with red lines. (C) Arrhythmia rate of the control (0.001% DMSO) and 24-hr terfenadine (20 μM)-treated zebrafish larvae (n=20 zebrafish/group). (D) Atrial size of the control (0.001% DMSO) and 24-hr terfenadine (20 μM)-treated zebrafish larvae (n=20 zebrafish/group).
A = atrium; DMSO = dimethyl sulfoxide; dpf = days post fertilization; FV/FA = fetal ventricle/fatal atrium; V = ventricle.
*p<0.050.

Transient terfenadine treatment impaired cardiac contraction, resulting in HF
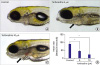
In terfenadine-treated zebrafish, blood frequently pooled and clotted in the chamber, and circulation was remarkably reduced (Figure 4A). Both atrium (white circle) and ventricle (red circle) were swollen, and fluid was deposited around the heart, mimicking edema (blue circle) (Figure 4B). Cardiac contractility was significantly reduced and ventricular area was significantly enlarged by terfenadine treatment (Figure 4C). The terfenadine-induced cardiac defects in zebrafish showed significant similarities with human failing hearts.
Figure 4
Transient terfenadine treatment impaired cardiac contraction, resulting in HF. (A) Representative images of control (0.001% DMSO) or terfenadine (20 μM)-treated zebrafish larva. (B) Lateral view of zebrafish larvae at 4 dpf. The control zebrafish (0.001% DMSO) exhibited normal cardiac morphology, whereas terfenadine (20 μM)-treated zebrafish larvae showed pericardial edema (blue circle) and venous congestion (green circle). (C) Quantification of ventricular FS in control (0.001% DMSO) and terfenadine (20 μM)-treated zebrafish larvae. Ventricle size of control (0.001% DMSO) and terfenadine (20 μM)-treated zebrafish larvae after a 24 hours treatment. n=20 zebrafish/group, scale bar=0.5 mm.
a = Atrium; DMSO = dimethyl sulfoxide; dpf = days post fertilization; FS = fractional shortening; HF = heart failure; V = ventricle.
*p<0.050; †p<0.010.

Terfenadine treatment induced cardiomyocyte death
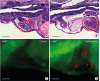
Next, we evaluated cardiomyocytes after terfenadine treatment. Myocardial cell apoptosis was evaluated by acridine orange staining. Apoptotic cells were identified with a florescence microscope as obvious bright spots (red arrow). There were no significant changes in cardiomyocytes treated by terfenadine on light microscopy with H&E staining (Figure 5B). However, as shown in Figure 5D, several terfenadine-treated cardiomyocytes showed apoptotic changes, which might reduce ventricular contraction.
Figure 5
Terfenadine treatment induced apoptosis in cardiomyocytes. (A) H&E staining of longitudinal sections showed normal cardiomyocyte morphology of control (0.001% DMSO) zebrafish larvae. (B) Terfenadine (20 μM)-treated zebrafish larvae for 24 hours showed elongated and thin cardiomyocytes. (C) Detection of apoptotic cardiomyocytes by acridine orange staining in the control group. (D) Detection of apoptotic cardiomyocytes by acridine orange staining in terfenadine-treated zebrafish. Outlines of ventricles are shown with red lines. Apoptotic cells are indicated by red arrows. Scale bar=0.5 μm.
DMSO = dimethyl sulfoxide; dpf = days post fertilization; H&E = hematoxylin and eosin.
Terfenadine treatment induced HF and impaired survival in zebrafish larvae
Lastly, we evaluated natriuretic peptide expression following terfenadine treatment. A significant increase of natriuretic peptide B (NPPB) mRNA was found in terfenadine-treated zebrafish larvae at 12 hours post-treatment. Short duration terfenadine significantly impaired survival in zebrafish. Low-dose terfenadine (5–10 µM) did not show mortality in short-term treatment (24 hours). However, moderate-dose (35–45 µM) terfenadine treatment reduced zebrafish survival within 1 hour, which suggested that cumulative treatment of terfenadine might cause HF-related death in zebrafish (Figure 6).
Figure 6
Terfenadine treatment induced HF and impaired survival in zebrafish larvae. (A) Quantitative RT-PCR of NPPB was increased in terfenadine-treated zebrafish larvae. (B) Survival rate of zebrafish treated with control and 35 to 45 µM of terfenadine. n=20 zebrafish/group.
HF = heart failure; NPPB = natriuretic peptide B; RT-PCR = real-time polymerase chain reaction.
*p<0.050.

DISCUSSION
HF is a progressive condition in which the heart muscle is unable to pump enough blood to meet the body's needs for blood and oxygen. We developed a zebrafish-dilated cardiomyopathy model using short-term terfenadine treatment.
We used terfenadine to induce QT prolongation and cardiac arrhythmia in zebrafish. Terfenadine-treated zebrafish showed enlarged atrial and ventricular size and venous congestion. Heart rate was markedly reduced even after terfenadine withdrawal. Compared with the control, terfenadine-treated zebrafish showed uncoupled atrial-ventricular contraction, demonstrating substantial blockage of atrioventricular conduction. Terfenadine-treated cardiomyocytes showed apoptotic change, reducing ventricular contraction. Significant increase of NPPB mRNA was found in terfenadine-treated zebrafish larvae at 12 hours post-treatment. Short-duration treatment with terfenadine significantly impaired survival in zebrafish.
The important finding of our study is that short-term terfenadine treatment can induce non-ischemic HF in zebrafish. Changes with terfenadine treatment in zebrafish were as follows. First, atrioventricular dyssynchrony and cardiac chamber enlargement were caused by a short duration of terfenadine treatment. Second, cardiac contractility was significantly reduced in terfenadine-treated zebrafish. Third, increased cardiomyocyte apoptosis and a significant increase of NPPB were observed in terfenadine-treated zebrafish. Lastly, moderate-dose terfenadine markedly impaired zebrafish survival.
Terfenadine is an antihistamine formerly used for the treatment of allergic conditions. However, it was superseded by fexofenadine in the 1990s due to the risk of cardiac arrhythmia caused by QT interval prolongation.1) In this study, we treated 4 dpf zebrafish with terfenadine for 2 hours to induce atrioventricular dyssynchrony, chamber enlargement, and pericardiac edema, which are prototypical responses in HF. The idea of using terfenadine came from a previous study by Tang et al.1) We evaluated the mechanism, phenotypic change, and HF development time frame.
Zebrafish has been a model for human cardiac disease for a much shorter time period than most other animal models, which helps explain the smaller number of total proteomic and cardiac related publications.23) Zebrafish have been used successfully in drug discovery and chemical screening processes.24) The growth and development of zebrafish embryo are very rapid, such that within 2 to 3 days, the larvae hatches from its chorion and has to survive as a free-swimming organism.24) Zebrafish embryos in the first few days of life are 1 to 3 mm in length and are suited for arraying into 96- and even 384-well plates, which are suitable for mass screening. High throughput chemical screens using zebrafish were first published in 2000 to analyze the developmental effects of structurally diverse chemical compounds in embryos.25) Using a 96-well plate, embryos were arrayed in each well, and the phenotypic effects of a compound were documented by visual inspection and included specific cardiac, brain, and growth defects. Beyond the benefits in mass screening, zebrafish models are useful in other aspects of cardiovascular disease research. The zebrafish heart consists of 2 chambers, one atrium and one ventricle, consisting of both a myocardium and an endocardium.26) In addition, the zebrafish is a vertebrate, and most of its genes have human homologs.23) Using these benefits of a zebrafish model, rare genetic diseases and chemotherapy-related cardiotoxicity have been investigated.27)28)
The main purpose of this study was to establish a zebrafish HF model. Using short-term treatment of terfenadine and cmlc-2:GFP zebrafish, we clearly showed cardiac chamber enlargement, depressed systolic function, and rhythm disturbance. Moreover, due to the genetic homology of zebrafish with human genes, NPPB change could also be evaluated. We evaluated the protective role of HF medication in a zebrafish HF model. We hoped to broaden our HF knowledge using a zebrafish HF model with mass screening method. The limitation of this study is the relatively short duration of observation with a limited dose range of terfenadine. With treatment of a given dose of terfenadine, HF physiology was maintained over 4 days. However, with variable doses of terfenadine, especially lower doses, there is the possibility that transient HF might be reversed spontaneously. Therefore, in future research evaluating the protective role of certain medications, confirmation of HF feature maintenance throughout the study period is necessary.
In conclusion, we induced secondary dilated cardiomyopathy by atrioventricular dyssynchrony with short-term terfenadine treatment. With the advantages of rapid sample preparation procedure and data analysis, a zebrafish HF model can potentially be applied to drug screening and toxicity assays for non-ischemic cardiomyopathy.
 XML Download
XML Download