

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Early reperfusion in acute myocardial infarction (AMI) is essential to limit infarct size. However, reperfusion can paradoxically induce a lethal ischemia/reperfusion (IR) injury.1) Ischemic preconditioning (IPC) and ischemic postconditioning (IPostC) utilize transient non-lethal episodes of myocardial IR either before or after the index ischemic event, respectively, and have been introduced as methods to reduce IR injury.2)3) Unfortunately, these techniques are not clinically relevant. Another strategy for reducing IR injury is remote ischemic conditioning (RIC). In brief, RIC involves reversible episodes of ischemia and reperfusion in a vascular bed, tissue, or organ, aimed at conferring a global protective phenotype, thereby rendering remote tissues and organs resistant to future IR injury.4) Strong evidence from both experimental and clinical studies supports the protective effect of RIC on IR injury in the heart as well as other organs. The detailed mechanisms involved in the local release of protective signals at remote sites and the subsequent contributions of neuronal and humoral pathways are not yet clear, not only in regard to signal release, but also the manner by which these signals are transferred to the target organ to mediate protective effects.
Classic experimental protocols for RIC in rats have utilized models consisting of either an elastic rubber band or tourniquet placed around a hindlimb to occlude arterial blood flow or a model of coronary effluent collected from an isolated donor heart after IPC.5)6)7)8) Considering the analysis of humoral factors as signal transfers, dialysate from the venous blood plasma vein in the tourniquet model is necessary, but this dialysate can be affected by unknown humoral factors present in systemic circulation. In contrast, signals from the coronary effluent collected from the heart might be different from those of distant sites such as the hindlimb. Thus, in order to facilitate analysis of humoral factors, there is a need for a new experimental model in which effluent is collected directly from the hindlimb under conditions where it is not affected by systemic circulation.
Apart from mechanisms of signal transduction, the specific cardioprotective signaling pathways activated in the target myocardium in RIC remain unclear. It has been shown that protection against IR injury produced by either IPC or IPostC involves activation of the reperfusion injury salvage kinase (RISK) pathway, among which phosphatidylinositol-4,5-bisphosphate 3-kinase, protein kinase B (Akt), and extracellular signal-regulated kinase (ERK) 1/2 play important roles.9)10) In IPostC, signal transducer and activation of transcription-3 (STAT-3) plays an important role in reducing IR injury as part of the survivor activating factor enhancement (SAFE) pathway.11)12) Recent data also suggests that the RISK and SAFE pathways likely interact in mediating RIC.5)6)7)8)13)14)15) Thus, in order to test whether the RISK and SAFE pathways were activated in our new RIC model, we assessed ERK 1/2 and STAT-3 phosphorylation status by immunoblotting.
The aims of the present study were 1) to examine whether the effluent from a novel isolated hindlimb model after RIC could reduce IR injury in an isolated heart model, 2) to examine the effect of effluent from the new isolated hindlimb model without RIC on infarct size in an isolated heart model, and 3) to clarify the involvement of the RISK and SAFE pathways during RIC.
MATERIALS AND METHODS
Animals and ethical statement
Male Sprague-Dawley rats were fed a standard diet at a central animal unit and were used at a weight of 250–350 g throughout the study. The experimental protocol was approved by the Institutional Animal Care and Use Committee in Yonsei University Wonju College of Medicine (YWC-150129-1).
Isolated hindlimb model for RIC
For the isolated hindlimb model, a rat was anesthetized via intraperitoneal (IP) administration of ketamine (60 mg/kg). An additional 1/3 to 1/2 dose of ketamine was given every 15 minutes as needed for effective anesthesia. The rat was tracheotomized and ventilated with room air supplemented with oxygen at 65 breaths/min. Atelectasis was prevented by maintaining a positive end-expiratory pressure of 5–10 cm H2O. Body temperature was maintained at 37°C using a heating pad. The schema of the perfusion system for the RIC model is presented in Figure 1A. The abdomen was opened in the median line, and the small and large bowels were moved to the right side of the abdomen. After exposing the abdominal aorta, the infrarenal aorta and inferior vena cava (IVC) were cannulated with 26-gauge plastic needles, which were advanced to the right common iliac artery (CIA) and common iliac vein (CIV), respectively. The aorta and IVC above the cannulated site, right CIA and CIV above the needle tips, and left CIA and CIV were tied with 5-0 silk to isolate right hindlimb circulation from systemic circulation. A dose of 200 IU of heparin was given via venous cannulation. Krebs-Henseleit buffer (KHB) from a Langendorff apparatus was perfused through the arterial cannulation, and effluent from venous cannulation was collected. The composition of KHB (pH 7.4, oxygenated with 95% O2/5% CO2) in mM was as follows: 118 NaCl, 4.7 KCl, 1.8 CaCl2, 1.2 H2PO4, 1.2 MgSO4, 25.2 NaHCO3, and 11.0 glucose. The perfusion pressure was maintained between 60 to 80 mmHg using a peristaltic pump. The KHB temperature was kept at a constant 37.0°C using a heat exchanger. After stabilization for 10 minutes, either the sham or RIC procedure was started (Figure 2A). The hindlimb effluent during stabilization was discarded because of blood. During the RIC procedure, ischemia was achieved by stopping the peristaltic pump for 5 minutes, which was followed by reperfusion for 5 minutes. The IR sequence of the RIC procedure was repeated 3 times, during which approximately 150 mL of hindlimb effluent was collected. In the sham procedure, approximately 150 mL of hindlimb effluent was collected for 15 minutes without interrupting the hindlimb perfusion. The effluent was maintained at room temperature during collection and was immediately used in the isolated heart model for IR injury.
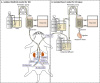 | Figure 1Schema of the perfusion system for the RIC model (A) and isolated heart model for IR injury (B). (A) After exposing the abdominal aorta, the infrarenal aorta and IVC were cannulated with 26-gauge plastic needles that were advanced to the right CIA and CIV, respectively. The aorta and IVC above the cannulated site, right CIA and right CIV above the tip of the needles, and left CIA and left CIV were tied with 5-0 silk to isolate right hindlimb circulation from systemic circulation. Heparin (200 IU) was given via venous cannulation. KHB was perfused through the arterial cannulation, and effluent from venous cannulation was collected. (B) The heart was rapidly excised via clamshell thoracotomy and placed in ice-cold KHB before being mounted on a Langendorff apparatus. KHB was used for retrograde perfusion at a constant pressure (60–80 mmHg). The KHB temperature was kept at a constant 37.0°C using a heat exchanger. A water-filled latex balloon, connected to a hydrostatic pressure transducer and coupled to a high-performance data acquisition system, was inserted into the LV and inflated to an end-diastolic pressure of 5–15 mmHg. CF was measured by timed collection of effluent over 1 minute.
CF = coronary flow; CIA = common iliac artery; CIV = common iliac vein; IR = ischemia/reperfusion; IVC = inferior vena cava; KHB = Krebs-Henseleit buffer; LV = left ventricle; RIC = remote ischemic conditioning; Rt = right.
|
 | Figure 2Design of experimental protocols. (A) RIC procedures. Anesthetized rats were subjected to either 1) a sham procedure, or 2) remote ischemia (3×5 minutes hindlimb IR using a Langendorff system). During the procedures, effluents were collected. (B) The heart from another rat was excised and perfused on a Langendorff system before being subjected to the IR protocol (30-minute regional ischemia and 60-minute reperfusion): 1) control group, 2) IPC group: prior to the IR protocol, IPC (3×1 minute LAD IR) was applied, 3) sham effluent group: effluent after sham procedure was perfused for 10 minutes before the IR protocol, and 4) RIC effluent group: effluent after RIC procedure was perfused for 10 minutes before the IR protocol. Following each Langendorff experiment, infarct size was determined using TTC staining.
IPC = ischemic preconditioning; IR = ischemia/reperfusion; LAD = left anterior descending coronary artery; RIC = remote ischemic conditioning; TTC = triphenyl-tetrazolium-chloride.
|
Isolated heart model for IR injury
The schema of the perfusion system used for the isolated heart model is presented in Figure 1B. During the isolated hindlimb model procedure, a separate rat was anesthetized via IP administration of ketamine (60 mg/kg) and heparinized (200 IU). The heart was then rapidly excised via a clamshell thoracotomy and placed in ice-cold KHB before being mounted on a separate Langendorff apparatus.16)17) Excised hearts were used only when the duration between thoracotomy and mounting was less than 3 minutes. KHB was used for retrograde perfusion at a constant pressure of 60 to 80 mmHg and was maintained at a constant temperature of 37.0°C with a heat exchanger. Coronary flow (CF) was measured by timed collection of effluent over 1 minute. The heart was stabilized for 10–20 minutes according to the experimental protocol. A 3-0 silk suture was passed around the left coronary artery and threaded through a small vinyl tube to form a snare. Regional ischemia was achieved by pulling the snare, which was confirmed by the presence of a pale area below the snare and a substantial decrease in CF. The IR protocol consisted of 30 minutes of ischemia and 60 minutes of reperfusion (Figure 2B). Although the duration of reperfusion can influence infarct size in the mouse Langendorff model, a 60-minute reperfusion duration in the rat Langendorff model does not influence the efficacy of triphenyl-tetrazolium-chloride (TTC) staining compared with a reperfusion period of 120 minutes.18)
Experimental protocols
As shown in Figure 2B, rats were randomly assigned to one of 4 groups: control group (IR protocol only, n=3), IPC group (3 cycles of 1 minute ischemia followed by 1 minute reperfusion prior to IR protocol, n=3), sham effluent group (10 minutes perfusion with hindlimb effluent after sham procedure prior to IR protocol, n=3), and RIC effluent group (10 minutes perfusion with hindlimb effluent after the RIC procedure prior to IR protocol, n=3).
Determination of area at risk (AAR) and infarct size
At the end of each experiment, the silk suture was securely tightened, and a 0.2% Evans Blue solution was infused to visualize the risk zone. The heart was then cut into 2-mm-thick slices parallel to the atrioventricular groove and stained with 1% TTC in phosphate buffer (pH 7.4) for 20 minutes at 37°C. Next, the slices were immersed in 4% formalin to enhance the contrast of the stain. Photographs of slices were taken by digital microscope. The left ventricle (LV) area, AAR, and infarct area (IA) were gauged on one side of a single slice using a computerized planimetry program. The LV area, AAR, and IA from each slice were then summarized. AAR was defined as the area not stained by Evans Blue and was presented as the AAR/LV area (%). IA was defined as the pale area not stained red by TTC and was presented as the IA/AAR (%).
Immunoblotting of survival kinases
Separate groups of rats (n=3 per group) were used for analysis of protein phosphorylation. Samples for immunoblotting were harvested after reperfusion for 15 minutes. Myocardium from the AAR was excised and processed with a snap freeze-clamp in liquid nitrogen prior to being stored at −80°C. In addition, frozen myocardial tissue samples were powdered in a mortar and pestle pre-cooled with liquid nitrogen. Approximately 100 mg of powdered myocardial tissue was used for protein extraction. Frozen myocardial tissue samples were homogenized on ice in 0.5 mL ice-cold lysis buffer containing 30 mM HEPES, 20 mM KCl, 2.5 mM EGTA, 2.5 mM EDTA, 40 mM sodium fluoride, 4 mM sodium pyrophosphate, 1 mM sodium orthovanadate, 10% glycerol, 1% Nonidet P-40, a phosphatase inhibitor cocktail (Sigma-Aldrich, St. Louis, MO, USA), and a protease inhibitor cocktail (Complete Mini; Roche Applied Science, Mannheim, Germany). The homogenate was centrifuged at 13,000 rpm at 4°C for 1 hour, and the supernatant was collected. Protein concentration was determined using a Bio-Rad DC protein assay kit (Bio-Rad, Hercules, CA, USA) according to the manufacturer's instructions. Aliquots of the supernatant containing equal amounts of proteins (40 μg) were heated to 95°C for 5 minutes in sample loading buffer. Proteins were separated on 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) gels and transferred to a polyvinylidene fluoride (PVDF) membrane (Millipore, Billerica, MA, USA). After blocking nonspecific binding sites with 5% nonfat milk for 1 hour in Tris-buffered saline with Tween (TBST) containing 20 mM Tris-HCl, 137 mM NaCl (pH 7.6), and 0.1% Tween-20, the membranes were incubated overnight at 4°C with rabbit antibodies against phospho-ERK 1/2, total ERK 1/2, 705Tyrphospho-STAT-3, or total STAT-3 (Cell Signaling Technology, Inc., Danvers, MA, USA). Beta-actin (Sigma-Aldrich) was used as a loading control. After washing in TBST, the membranes were incubated for 1 hour at room temperature with horseradish peroxidase conjugated anti-rabbit immunoglobulin G (IgG) secondary antibody (1/2,000; Santa Cruz Biotechnologies, Santa Cruz, CA, USA), and the bound antibody was detected using an enhanced West Pico chemiluminescent substrate (Thermo Fisher Scientific, Rockford, IL, USA). Band densities of the appropriate molecular mass (42/44 kDa for ERK 1/2, 79/86 kDa for STAT-3) were determined semi-quantitatively using a biospectrum 600 imaging system (UVP Inc., Upland, CA, USA).
Statistical analysis
All values are presented as the mean±standard error of the mean (SEM). Statistical analyses were performed using SPSS Statistics 20.0 software (IBM, Armonk, NY, USA). Differences between groups were evaluated using one-way analysis of variance (ANOVA) combined with post hoc Fisher's least significant difference (LSD)-corrected multiple comparison test. A value of p<0.050 was considered statistically significant.
RESULTS
The isolated hindlimb model was successfully applied to a total of 12 of 15 (80%) rats. There were 3 cases of technical failure involving perforation or rupture of a vessel. Likewise, the isolated heart model was successfully applied to a total of 24 of 32 (75%) rats. Rats were excluded from data analysis if they could not continue the procedure due to unstable hemodynamics such as sustained ventricular fibrillation (n=4) or asystole (n=2) or if there was a technical failure such as contamination by Evans Blue solution (n=2).
Effluent from the isolated hindlimb model after RIC induces significant cardioprotection
Representative sections of TTC-stained hearts following the IR protocol in each group are presented in Figure 3A. The AAR was not different among groups (Figure 3B). Effluent from the isolated hindlimb model after RIC was effective at inducing cardioprotection, as demonstrated by a significant reduction in IA compared to hearts subjected to effluent from the control group (31.2%±3.8% in control group vs. 20.6%±1.8% in RIC effluent group, p<0.050; Figure 3C). Compared with the IPC group, the RIC effluent group showed a similar but slightly lower degree of cardioprotection in terms of a reduction in IA (16.0%±3.9% in IPC group vs. 20.6%±1.8% in RIC effluent group, p=0.220; Figure 3C).
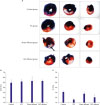 | Figure 3Representative sections of TTC-stained hearts following ischemia for 30-minute and reperfusion for 60-minute. (A) Bar graph showing the AAR expressed as a percentage of total left ventricular area (B) and IA as a percentage of AAR (C). All data is expressed as mean±SEM. n=3 in each group.
AAR = area at risk; IA = infarct area; IPC = ischemic preconditioning; RIC = remote ischemic conditioning; SEM = standard error of mean; TTC = triphenyl-tetrazolium-chloride.
*p<0.050 vs. control group.
|
Sham effluent from the isolated hindlimb model does not affect infarct size
The effluent from the isolated hindlimb model without RIC did not reduce IA compared to the control (31.2%±3.8% in control group vs. 27.7%±6.2% in the sham effluent group, p=0.330; Figure 3C).
Effluent from the isolated hindlimb model after RIC activates the SAFE pathway but not the RISK pathway
Western blot results of ERK 1/2 and STAT-3 phosphorylation in rat hearts after 30 minutes of ischemia followed by reperfusion for 15 minutes are presented in Figure 4. Compared with the control group, phosphorylation of ERK 1/2 was significantly increased only in the IPC group (p<0.050). Phosphorylation of STAT-3 showed a similar pattern to that of ERK 1/2 phosphorylation; however, phosphorylation of STAT-3 was significantly enhanced in all 3 groups compared with the control group (all p<0.050). Compared with the IPC group, the RIC effluent groups showed a similar increase in phosphorylation of STAT-3, while the sham effluent group had significantly lower phosphorylation of STAT-3.
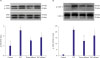 | Figure 4Western blot analysis of ERK 1/2 (A) and STAT-3 (B) phosphorylation in rat hearts subjected to ischemia-reperfusion. Top: representative immunoblots of p-ERK 1/2 and total ERK 1/2 (A) and p-STAT-3 and total STAT-3 (B) in LV homogenates from hearts subjected to ischemia-reperfusion. Bottom: bar graphs show mean±SEM of the densitometry of p-ERK to ERK ratio (A) and p-STAT-3 to STAT-3 ratio (B). Data are expressed as the mean±SEM. n=3 in each group.
AU = arbitrary units; ERK = extracellular signal-regulated kinase; IPC = ischemic preconditioning; LV = left ventricle; p-ERK = phosphorylated ERK; p-STAT-3 = phosphorylated STAT-3; RIC = remote ischemic conditioning; SEM = standard error of mean; STAT-3 = signal transducer and activation of transcription-3.
*p<0.050 vs. control, †p<0.050 vs. IPC.
|
DISCUSSION
To the best of our knowledge, this is the first study to investigate the effect of effluent from an isolated hindlimb on IR injury in an isolated heart model. In this study, we demonstrated that effluent after RIC was effective in decreasing IR injury in an isolated rat heart model. Consistently, we found that effluent obtained from a sham hindlimb model had no effect on infarct size. Finally, we demonstrated that the cardioprotective effects of effluent after RIC-involved recruitment of the SAFE pathway.
New model for RIC experiments: Following the first description of remote preconditioning by Przyklenk et al.19) in 1993, subsequent studies have demonstrated that IR injury can be reduced by RIC of remote organs such as the skin, mesentery, kidney, and limbs not only from brief local IR, but also from surgical trauma, autacoids, and possibly peripheral nervous stimulation.4) Identification of the cardioprotective factors generated by RIC has significant potential for the development of pharmacological therapies that can protect against IR injury after an AMI. Indeed, there have been several efforts to elucidate the detailed mechanisms behind local release of protective signals from remote sites as well as the contributions of neuronal and humoral pathways toward signal release and subsequent transduction of these signals to target organs. With respect to humoral signal transfer from remote organs to the heart, Dickson et al.20) demonstrated that the effluent collected from isolated rabbit donor hearts during IPC can reduce the infarct size from the IR injury in isolated recipient rabbit hearts. Specific to rats, studies by Serejo et al.,21) Breivik et al.,8) and Huffman et al.7) demonstrated the beneficial effect of effluent collected from a donor heart during IPC, as evidenced by reduced infarct size and improved myocardial performance in isolated recipient hearts. Using a fractionation approach, Breivik et al.8) showed that cardioprotective humoral factors include molecules that are hydrophobic in nature and less than 30 kDa, while Serejo et al.21) showed that the cardioprotective factors comprise thermolabile hydrophobic substances with molecular weights greater than 3.5 kDa. Although such analyses of effluent are important with respect to elucidating mechanisms of signal transduction, the effluents used in the aforementioned studies originated from the heart rather than from remote sites such as the limbs.
Most clinical studies on RIC in cardiac surgery, elective percutaneous coronary intervention (PCI), and AMI have utilized an RIC regimen involving the upper or lower limb. Thus, analysis of effluent from a limb, rather than from the heart, should provide more clinically relevant information regarding signal transduction. Recently, Skyschally et al.13) showed that the dialysate from plasma sampled after RIC in a pig hindlimb reduces IR injury in an isolated rat heart. Likewise, Pickard et al.15) demonstrated a similar cardioprotective effect of dialysate collected from a rat after tourniquet-mediated hindlimb RIC. Although analysis of dialysate from blood sampling after RIC in a hindlimb represents a good methodologic approach for elucidating the exact mechanisms of RIC in the remote organ, such methods require complex processing and expensive equipment. In addition, the dialysate might be affected by unknown humoral factors from the systemic circulation; thus, any intervention during RIC might limit the applicability of this approach.
The most appealing aspects of the isolated heart model are its ability to facilitate study of the effects of interventions and avoidance of extrinsic factors that could influence study outcomes. For example, Weinbrenner et al.22) developed a rat infra-aortic occlusion model to test the effect of RIC effect in vivo.22) Specifically, they developed this model after determining that RIC through mesenteric or renal ischemia as a protective trigger is not effective in rats, and that ligation of the femoral artery or limb ischemia induced by placing a thin elastic tourniquet around the hindlimb is similarly ineffective. Several different investigators have found that the rat infra-aortic occlusion model is both reliable and reproducible and lends itself to the investigation of the protecting signals involved with remote preconditioning, especially since the venous blood of the hindlimb is easily collected by puncture of the IVC. Based on the model developed by Weinbrenner et al.,22) we designed an isolated hindlimb model similar to the isolated heart model. Using this model, we showed that effluent obtained after RIC had significant cardioprotective effects, whereas effluent after a sham procedure did not affect infarct size. Although we did not analyze the effluent composition as part of this study, we expect that our model will assist investigators in clarifying the protective mechanisms of RIC.
Involvement of the RISK pathway: Our results showed that phosphorylation of ERK 1/2 was significantly increased in the IPC group, but not in the RIC effluent group. Conversely, the level of ERK 1/2 phosphorylation was higher in the RIC effluent group than the control group, although the difference did not reach the level of statistical significance. Several previous studies have shown that RIC results in activation of the RISK pathway, as demonstrated by activation of Akt and ERK 1/2.5)6)8)13) Likewise, inhibiting RISK activation by pharmacological antagonist exacerbates IR injury. For example, Breivik et al.8) demonstrated that effluent collected from isolated rat hearts during IPC, when administered to a naïve recipient rat heart as an RIC stimulus, reduces IR injury through phosphorylation of phosphatidylinositol 3-kinase (PI3K)-Akt. They also showed that the cardioprotection achieved with the effluent was abolished when a PI3K inhibitor and Akt-inhibitor were co-administered during reperfusion with effluent. Similarly, Xin et al.5) showed that RIC consisting of 4 cycles of 5 minutes of limb ischemia/5-minute reperfusion with occlusion of blood flow to one hindlimb by a tourniquet reduces IR injury via the PI3K/Akt and ERK 1/2 pathways, and that this cardioprotection is abolished by administering a PI3K-selective inhibitor and ERK 1/2 inhibitor. In that study, phosphorylation of Akt and ERK 1/2 was significantly higher in the RIC group compared with the control group, but was significantly lower in the RIC group compared with the IPC group, which is consistent with the results of the present study. Xin et al.5) also showed that the combination of IPostC and RIC reduces infarct size in IR injury compared to IPostC alone or RIC alone and also that the degree of phosphorylation of Akt and ERK correlates with infarct size. Thus, the lack of a statistically significant increase in the phosphorylation of ERK 1/2 in the RIC effluent group in our study might be related to a dose-dependence of ischemic conditioning. On the other hand, because the pattern of ERK 1/2 phosphorylation was similar to that of STAT-3 phosphorylation in our study, future studies utilizing increased animal numbers, a concentrated effluent, different RIC protocol, or a more sensitive antibody assay might identify a significant increase in phosphorylation of ERK 1/2. Thus, while the results of our study did not exclude the possibility of RISK pathway activation, they also did not confirm RISK pathway activation. In further support of the possibility of RISK pathway activation as a mechanism of RIC, Heidbreder et al.14) demonstrated that cardioprotection by RIC induced by occlusion of the mesenteric artery results in activated mitogen-activated protein kinases (MAPKs) such as p38, ERK 1/2, and c-Jun N-terminal protein kinase (JNK) 1/2 within the intestinal tissue but not within the myocardium.23)
Involvement of the SAFE pathway: Phosphorylation of STAT-3 was significantly increased in the IPC, RIC effluent, and sham effluent groups, and the increase was greatest in the IPC and RIC effluent groups. STAT-3 is a central element of the SAFE pathway, and Huffman et al.7) demonstrated that effluent collected from the isolated rat heart models during IPC improves myocardial performance by activating Janus kinase-signal transducer and activator of transcription (JAK-STAT) signaling. Tamareille et al.6) demonstrated that phosphorylation of STAT-3 is not significantly increased following either IPostC alone or RIC alone, whereas the combination of IPostC and RIC significantly increases phosphorylation of STAT-3. Finally, Skyschally et al.13) demonstrated that dialysate from plasma sampled after RIC in a pig hindlimb, but not in a rat hindlimb, reduces IR injury in isolated rat hearts in a process involving phosphorylation of STAT-3. Collectively, these results are consistent with the results of the present study. However, phosphorylation of STAT-3 was also significantly higher in a sham effluent group compared with the control group in our study, although this phosphorylation was significantly lower than that of the IPC or sham effluent groups. There is some evidence to suggest that the RIC effect can be achieved by surgical trauma through neuronal signaling mechanisms.24252627) Although we used an anesthetic agent before procedure, a surgical procedure is a necessary aspect of our isolated hindlimb model. Thus, the phosphorylation of STAT-3 and ERK 1/2 in the sham effluent group might have been due to a neuronal signaling mechanism caused by the surgery. Interestingly, infarct size was slightly smaller in the sham group compared with the control group.
Limitations: As described in the Discussion, we designed a new isolated hindlimb model aimed at minimizing the effect of systemic circulation apart from nervous system activation. Thus, the first limitation of this study was that, although we used anesthesia to reduce pain during the procedure and a considerable time of stabilization before collecting the effluent, the procedure itself involved surgical trauma. For this reason, activation of neural pathways could not be excluded as a mechanism to explain our results. Indeed, infarct size was slightly smaller in the sham effluent group compared to the control group, and phosphorylation of ERK 1/2 and STAT-3 was also increased in the sham effluent group. A second limitation of this study was that we used effluent obtained before starting ischemia in the IR model. Although we used KHB for the RIC model, the characteristics of KHB after RIC might have changed during processing, including a change in pH due to increased lactic acid levels or production of reactive oxygen species. Thus, the differences seen with IPC might have been due to differences in the KHB. The small sample size of each group was a third limitation of this study, and the problems associated with groups of small numbers of animals have been well described by Sachdeva et al.28) In small groups, the presence of 1 or 2 outliers can result in false positive or false negative results, with 1 or 2 missed coronary occlusions resulting in a smaller mean infarct size. Similarly, 1 or 2 rats with a coronary anatomy that lends itself to small infarcts such as higher collateral flow in the risk zone can lead to a smaller mean infarct size for the group. Thus, future studies with larger numbers of animals are needed to confirm our results.
In conclusion, we developed a new isolated hindlimb model for improved effluent collection. Using this model, we confirmed that RIC effluent could effectively reduce IR injury. In addition, we found that the cardioprotection by RIC effluent was associated with activation of the SAFE signaling pathway.
 XML Download
XML Download