

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Defects in cardiac development become the causes of miscarriage. More than 80% of early abortion is thought to be ascribed to the chromosome abnormality. Among the total pregnancy, 12-15% ends in miscarriage.1)2) However, even after the heart beat is clinically recognized, some embryos do not develop due to unknown causes. Severe congenital heart diseases (CHDs) might be the cause of cessation of pregnancy. One percent of new babies are estimated to have CHDs.3)4) Mutations of transcription factors including Nkx2.5, Tbx5, GATA family transcription factors, and MEF2, which regulate cardiogenesis, result in CHDs (atrial septal defect, ventricular septal defect, tetralogy of Fallot, double outlets of right ventricle, and transposition of great arteries).5)6) Besides the regulation of genetic program-dependent expression of transcription factors for mesodermal cells that give rise to cardiomyocytes, the defects in the connection of great vessels that also originate from the mesoderm along with the heart result in CHDs.7) These abnormal developments of the hearts and the vessels are thought to be ascribed to the abnormal left-right axis formation during early embryogenesis.
Zebrafish is a useful model to study embryogenesis, because the embryos of zebrafish are transparent, can be manipulated easily to delete genes of interests and introduce transgenes, and grow extra-maternally.8)9)10)11) Early cardiogenesis can be monitored by cardiomyocyte-specific promoter-dependent fluorescent protein expression. Zebrafish has single atrium and single ventricle, while mouse and human have four chambers. Although this structural difference is present between mammals and zebrafish, the most genetic transcriptional regulation is conserved in zebrafish and mammals.12)13) Therefore, the regions of promoter/enhancer of the transcription factors have been used for marking cardiomyocytes.14)15) In addition to visualizing cardiomyocytes, the endocardium consisting of endothelial cells is observed using endothelium-specific promoter-controlled fluorescent protein expression.16) Furthermore, beside these mesoderm-derived cells or precursor cells, endodermal cells can be simultaneously visualized using endoderm-specific promoter-regulated fluorescence. The imaging of the embryos using the combination of the distinct tissue- or organ-specific promoters and fluorescent proteins enables us to demonstrate how the endoderm and the mesoderm simultaneously develop with mutual effect on each other.
Initial body planning is determined mainly by three axes: rostral-caudal axis, dorsal-ventral axis, and left-right axis.17) Among them, the left-right axis determination defines the position of the heart. This left-right axis formation follows the Kupffer's vesicle (KV) formation that corresponds to the Node of mammals. The dorsal forerunner cells (DFCs) of the endoderm give rise to KV-forming cells.18)19) Subsequently, these cells become ciliated to promote unidirectional flow in the KV.20) Motile cilia rotate unidirectionally from right to left to induce unidirectional flow in the KV.21) This flow leads to the asymmetric expression of Nodal, Lefty, and Pitx2 in the left side of the embryo,22) which causes left-right asymmetric patterning, which determines the position and the fate of cardiac precursor cells (CPCs) present in the lateral plate mesoderm.
We introduce our recent findings about the CPC migration governed by the endodermal cells in zebrafish. Furthermore, we overview how left-right axis is determined for spatial patterning of the heart during early embryogenesis and focus on what will be clarified to understand left-right asymmetry of organ position.
Specification and Determination of CPCs at the Blastula Stage
Even at the blastula stage of zebrafish embryos, the localization of the cells that are going to differentiate into atrial and ventricular precursor cells is determined.23) This lineage tracing is performed by the caged fluorescein. By tracking the fluorescein-positive cells, the initially positioned cells are analyzed for their movement before and after gastrulation. These results suggest the regulatory mechanism underlying the specification and spatial positioning of CPCs during and after gastrulation. Retinoic acid, canonical Wnt, bone morphogenetic protein, Nodal, and fibroblast growth factor are reported to regulate the number of cardiomyocytes by directly acting on CPCs or by indirectly affecting the pattering of three germ layers and subsequent positioning of CPCs.24)25)26)27)28) These signaling patterns during blastula stage are nicely summarized by Schier and Talbot.17) The signaling molecules externally affecting CPCs determine the positioning of three germ layers that are required for the formation of dorsal-ventral and rostral-caudal axes. Apelin and its receptor (Agtrl1b)-mediated signal determines the anterior-posterior length during gastrulation, thereby affecting the positioning of the heart.29) Agtrl1b is also expressed in the CPCs in the lateral plate mesoderm. Therefore, Apelin might directly regulate the migration of CPCs towards the midline (as described below in “heart tube formation”). Another axis (left-right axis) determined by the KV-mediated left-right asymmetry governs the positioning of heart, because the CPCs are positioned in the bilateral anterior lateral plate mesoderm by the 6-9 somite stages (ss) when the KV is completely organized.
While axes are determined during the blastula stage and the gastrula stage, hierarchical transcription factor-dependent cardiomyocyte specification progresses in the mesoderm. Transcriptional activation in the other two layers might affect cardiogenesis from early stage (specification) to late stage (the migration of CPCs toward midline to form heart tube). A mutant of Gata5 (faust) exhibits endodermal defects that cause cardia bifida.30) That of Raldh2 (necklace) exhibits an increase in cardiomyocytes as well as spinal cord defects.31) The CPCs express Gata family transcription factors (Gata4, 5, and 6), Nkx2.5, Hand2, and Tbx5. However, it remains unclear how the expression of these transcriptional factors are precisely regulated during the blastula stage and the gastrula stage.
Contribution of Endodermal Cells in Determination of Left-Right Axis
Left-right asymmetry is determined by several steps (Fig. 1). The unidirectional flow by counterclockwise rotation of cilia in the KV of zebrafish is essential for left-right axis formation. Therefore, the establishment of KV formed by DFCs is of importance.32) However, it remains unclear the manner in which two dozens of DFCs are selected among endodermal cells and how these cells move posteriorly.19) The requirement of endodermal cells is proven by mutant fish embryos in which sox32 is depleted.33) A sox32-depleted mutant (Casanova) lacks the KV. Moreover, the essential role of DFCs in forming the KV is clearly demonstrated by the experiment in which DFCs are ablated.32) The T-box transcription factor, no tail, the nodal co-receptor, and one-eyed pinhead are also essential for formation of the KV, although these are not specifically expressed in the endoderm,32)34) suggesting mutual regulation among the ectoderm, endoderm, and mesoderm to form the KV.
DFCs can be visualized by the transgenic fish embryos, Tg (sox17: green fluorescent protein [GFP]),35) in which GFP expression is promoted by sox17 promoter activated in the endoderm (Fig. 2). Therefore, Tg fish can be employed to examine migration and morphology of DFCs and to inspect the effect of depleting genes of interests. The KV is formed during early stages (4-6 ss). When the DFCs migrate, they need to become a cluster and move in a manner dependent on collective cell migration. Thus, cell-cell adhesion tightened by Cadherin-1 is essential for migration.32) In later stage, the KV must form a rosette-like structure with a lumen filled with fluid. The migration of DFCs and the subsequent formation of cluster leading to the KV are disturbed by the reduction or the depletion of FGF signal regulators including Fibroblast growth factor receptor-1a (Fgfr1a), Fgf8, and Canopy1.36)37)
Kupffer's Vesicle for Determination of Left-Right Asymmetry
The KV is a transient structure formed during embryogenesis to determine left-right asymmetry.21) Defects in ciliogenesis and disruption of the KV lead to abnormal left-right patterning, suggesting the essential role of ciliated cells in the KV (Fig. 3). The reduction in molecules constituting intraflagellar transport machinery of cilia: Ift57 (hippi), Ift88 (polaris), and left-right dynein related (Lrdr1), results in the defects of ciliogenesis and subsequent disruption of left-right patterning (Fig. 1). Cilia-dependent unidirectional flow causes the laterality of expression of Southpaw (Spaw), a nodal-related protein, Lefty2, and Pitx2 in the left side of lateral plate mesoderm. The expression of these Nodal inhibitors is regulated by positive feedback loop of the Spaw. On the other hand, in the right side, the expression of Charon increases to suppress the Spaw-dependent transcription by binding to the Spaw, since the flow starts in the KV. However, it still remains controversial as how the flow in the KV triggers and initiates the expression of these genes.
There exist at least two models to initiate the left-right asymmetry by a unidirectional flow: one is morphogen gradient; the other is two cilia model in the node of mouse. In the mouse node, cilia-mediated flow promotes morphogen gradient between the left and right sides. The flowing particles are suggested to bring morphogens preferentially to the left side.38) Sonic Hedgehog and retinoic acids are thought to be morphogens to be delivered by the flow. In the latter model, rotating motile cilia promotes the flow, while the immotile cilia on the perinodal crown cells sense the flow. This is hypothesized from the fact that the PKD2 in mutant mice is expressed in the cilia of perinodal crown cells thus exhibiting the left-right asymmetry of organ positioning.39) Therefore, PKD2-dependent sensing mechanism in cilia is a key determinant for the left-right asymmetry. In the KV, there are no clear data on morphogens. Further studies are required to examine whether the sensing cilia are present in the KV.
Calcium ion (Ca2+) increases in the left side of the KV as compared to the mouse node.40)41) In the zebrafish, a Ca2+ indicator, flash-pericam, can monitor the increase in Ca2+ in the KV. Ca2+ increases in the left side at the 4-6 ss via a 1,3,4,5,6-pentakisphosphate 2-kinase,41) although how this inositol phosphatase activity is regulated in the KV. In addition, Ca2+ oscillation in the cilia is thought to regulate left-right asymmetry.42)43)44) Although the cilia of the KV are motile,20) they are proposed to express PKD2 and sense extracellular Ca2+, as the immotile cilia in mouse perinodal crown cells do, although the cilia have 9+2 type microtubules in the KV.4) The movement of cilia and intracellular Ca2+ can be monitored by the small GTPase, Arl13b, tagged with genetically encoded Ca2+ indicators, GCaMP5 or RGECO1.42) However, recently, this concept has been challenged by the data that primary cilia do not respond to mechanical stimuli.45) In the reported study, the dominant Ca2+ oscillation in the left side of the node was not observed in contrast to the previous work.40) Therefore, although Ca2+ increases in the KV at the 4-6 ss and is required for left-right asymmetry, it is still unclear how an increased Ca2+ in the KV could result in left-right asymmetry, which determines the position of organs including the heart.
Congenital Heart Diseases and Ciliopathy
The node of mice corresponding to the KV of zebrafish determines the left-right asymmetry by nodal flow similar to the flow of the KV. Thus, the defect in nodal cilia results in the misplacement of organs in the body. Mouse nodal cilia lack the central pair of microtubules although they have nine pairs of microtubules with dynein arms. Therefore, cilia are grouped into three classes; normal motile (9+2) cilia, nodal cilia (9+0), and nonmotile (primary or sensory) cilia (9+0).46) Mutations in molecules that constitute cilia lead to ciliopathies.47) Therefore, several organ abnormalities are found in the patients with cilia-related genes. One of the best studied among the ciliopathies is primary ciliary dyskinesia (PCD). About 50% of patients with PCD exhibit abnormal organ positioning; heterotaxy and situs inversus, while the rest show situs solitus.48) The situs inversus is ascribed to the abnormal function of nodal cilia. Kartagener syndrome belongs to PCD and is caused by mutation in DNAI1 and DNAH5.49) Mutations of other ciliopathy-related genes cause CHD, although the organ positioning seems to be situs solitus. Consistently, forward genetic screening of mice reveals that most of the CHD are associated with mutations of cilia-related genes.47)
Migration of Bilateral Myocardial Precursor Cell Towards Midline to Form a Cardiac Cone and Subsequent Tube Formation
Atrial and ventricular CPCs that might not be fully differentiated migrate towards the midline to form a cardiac cone.50) As described above, CPCs are specified and differentiated during the blastula stage and the gastrula stage. Even after receiving the signals from left-right patterning molecules, these myocardial precursor cells might undergo fully differentiation process to become mature working cardiomyocytes. Myocardial precursor cells that are located in the bilateral lateral plate mesoderm move between the endoderm and the yolk syncytial layer (YSL) and migrate towards the midline (Fig. 4). Therefore, the defect in the endoderm and the signaling from YSL affects the migration of CPCs. A mutant of endoderm-specific transcription factor, sox32 (casanova) exhibits endodermal defects. Therefore, the defect in endoderm results in the inhibition of CPC migration towards midline, which ultimately leads to cardia bifida (two hearts).33)51)
A mutation in fibronectin (natter) causes cardia bifida, suggesting the importance of extracellular matrix for CPC migration.52) Notably, two mutations in molecules that function in sphingosine-1-phosphate (S1P)-mediated signaling cause cardia bifida: one is mil in which S1P receptor 2 (S1P2) gene is mutated;53) and the other is spsn2 in which S1P transporter gene is mutated.54)55)
It is of note that cardiomyocytes of zebrafish originate from first heart field (FHF) and second heart field (SHF) similar to mammalians.56) The draculin (drl) promoter-activated lateral plate mesodermal cells constitute FHF-derived cardiomyocytes when cardiac tube forms. On the other hand, the latent Tgfβ-binding protein 3 (ltbp3) promoter-activated cells become SHF-derived cardiomyocytes.57) The SHF-derived cardiomyocytes are positive for a homeobox transcription factor, Islet-1, in mammals as well as zebrafish. Although Ltbp3-postive cardiomyocytes are present in the arterial pole of the heart, other ltbp3-positive cells become both smooth muscle cells and endothelial cells. Because the venous pole cells are thought to originate from the SHF as evidenced by the fact that they are positive for Islet-1, it should be clarified how Ltbp3 contributes to the differentiation of the SHF precursors to SHF-derived cardiomyocytes as well as how the Ltbp3-negative cells contribute to the SHF formation.
Proper Endoderm Formation is Required for Cardiac Precursor Cell Migration
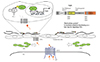
We have investigated as how S1P signaling regulates the migration of CPCs. Since we identified Spns2 as a S1P transporter functioning in the YSL, we had tried to investigate whether S1P-S1P2 signal is activated and found that the S1P released from the YSL activates S1P2 expressed in the endoderm.58) Spns2 mutant embryos showed the defect in the anterior endoderm where CPCs migrate towards the midline. Consistently, when S1P2 expression was reduced by antisense morpholino oligonucleotide, the morphant exhibited the endodermal defects similarly to the Spns2 mutant. We further demonstrated that the activation of S1P2-dependent signal induces nuclear translocation of Yap, a target of Hippo signaling. Therefore, we concluded that S1P released from the YSL activates S1P2 expressed in the endoderm, thereby inducing nuclear translocation of Yap to maintain the endodermal sheet that is required for CPC migration (Summarized in Fig. 5). Our results highlight the importance of the endoderm in a non-cell autonomous manner when the CPCs migrate towards the midline.
We have reviewed S1P-dependent signaling in the cardiogenesis of zebrafish. S1P-S1P receptor signaling remains elusive in cardiogenesis of mammals, although it has been extensively investigated in vascular development.59)60) Because cardiomyocytes, especially, trabecular cardiomyocytes express S1P1 receptors, cardiomyocyte-specific deletion of other S1P receptor genes will provide us with the information about S1P-dependent signaling in cardiogenesis.61)
As we described the importance of the endoderm when the KV forms and during both blastula and gastrula stages for the allocation and the specification of CPCs, the cardiac development is affected by the other tissues and organs. The signaling from the heart to other tissues might function vice-versa to precisely coordinate the whole body planning.

 XML Download
XML Download