

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Bone marrow-derived mesenchymal stem cells (BMCs) are multipotent cells and have been regarded as a promising regenerative cellular therapy in the treatment of ischemic diseases. BMCs are able to differentiate into vascular lineage cells and can be directly incorporated into newly formed vessels.1) Myocardial infarction remains a major cause of morbidity and mortality, and cell therapy has been aroused as a promising option for cardiac repair.
Our previous report showed that stem cells isolated from diabetic rats had a significant decline in cell proliferation and angiogenic activity, and oxytocin treatment restored the angiogenic function.2)
Diabetes, obesity, and aging are important stem cell microenvironments and may regulate stem cell function.3)4)5) In particular, diabetes is a well-known independent risk factor for cardiovascular disease, stroke, peripheral arterial disease, cardiomyopathy, and congestive heart failure,6)7)8)9) while diabetes is closely related with poor neovascularization after ischemia.10) Stem cell function to repair and support the injured myocardium declines with age or diabetes, which were representative limitations in the application of an autologous stem cell therapy for aged or diabetic patients with cardiovascular diseases. There is no doubt that diabetes is considerably associated with dysfunction of stem cells, and the responsible factors for diabetes-related dysfunction in stem cells need to be explored.
In this study, we compared the gene expression pattern of BMCs isolated from diabetic rats (DM-BMCs) with that of control cells (non-DM-BMCs), and suggested that Angiopoietin-like 4 (Angptl4) and micro ribonucleic acid-132 (miR-132) could be therapeutic targets for the repair of angiogenic function.
Materials and Methods
Experimental hyperglycemia
The study was approved by the Chonnam National University Institutional Animal Care and Use Committee (Chonnam National University AICUC-H-2010-11). Twelve-week-old adult male Sprague-Dawley rats (Central Lab Animal, Seoul, Korea) received intraperitoneal injections of streptozotocin (65 mg/kg, Sigma-Aldrich, St. Louis, USA) dissolved in 50 mM sodium citrate buffer (pH 4.5) to induce hyperglycemia. Age-matched controls were injected with an equivalent volume of saline. Fasting blood glucose levels were measured in tail veins, and rats with a blood glucose level >250 mg/dL were considered to be diabetic and were included in the study.
Cell cultures
Bone marrow cells were obtained from the tibia and femur under sterile conditions by using a syringe to flush the cavity out with warmed phosphate-buffered saline (PBS), collected by centrifugation, and resuspended with Dulbecco's Modified Eagle's Medium (DMEM) with 10% fetal bovine serum. Cells were plated into culture dishes, and non-adherent cells were removed by changing the medium after 72 hours. All cells used in this study were from the third and fourth passages of BMCs. Oxytocin (100 nM, Sigma-Aldrich, St. Louis, USA) was added to the growth medium for 24 hours for the experiments.
Human BMCs (hBMCs) immortalized by introduction of telomerase were kindly provided by Professor Yeon-Soo Kim (Inje University, Seoul, Korea). High glucose stress was induced by addition of 25 mM of glucose while 25mM of mannose was used as osmotic control.
In vitro angiogenesis assay
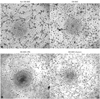
Tube formation was assayed by using an in vitro angiogenesis assay kit (Chemicon, Billerica, USA). Cells (1×104) were plated onto matrix gel-coated 96-well plates and were cultured in DMEM without serum. Tube formation was monitored and photographed by using an inverted microscope (Olympus, Tokyo, Japan), and images were analyzed by using Image-Pro software (MediaCybernetics, Rockville, USA). Angiogenic activity was quantified by measuring tube length and tube area. Total tube length in four fields per well was averaged, and three wells were used to produce one value per condition.
Microarray analysis
To identify the high glucose-responsible genes, we performed complementary deoxyribonucleic acid (cDNA) microarray using DM-BMCs and non-DM-BMCs with or without oxytocin treatment (GenomicTree, Daejeon, Korea). Ribonucleic acid (RNA) quality was assessed using the Agilent 2100 Bioanalyzer, and RNA was amplified by Agilent's Low RNA Input Linear Amplification kit PLUS and hybridized to Agilent Rat expression 4X44K (v3).
Reverse transcriptase-polymerase chain reaction and quantitative real-time polymerase chain reaction
To compare the messenger RNA (mRNA) expression level of Angptl4, cells were homogenized in Trizol (Invitrogen, Carlsbad, USA) according to the manufacturer's instructions. cDNA was generated by use of MMLV transcriptase (Invitrogen, Carlsbad, USA). Real time polymerase chain reaction (PCR) was performed using a QuantiTect SYBR Green PCR kit (Qiagen, Valencia, USA) and Corbett Research Rotor-Gene RG-3000 Real Time PCR System. Primers were purchased (Bioneer, Daejeon, Korea).
Western blot analysis and enzyme-linked immunosorbent assay
Cells were resuspended in lysis buffer (20 mM Tris-HCl, pH 7.4, 0.1 mM ethylenediaminetetraacetic acid, 150 mM NaCl, 1 mM phenylmethylsulfonyl fluoride, 1 mg/mL leupeptin, 1 mM Na3VO4), and kept on ice with occasional tapping for 20 minutes. After centrifugation at 10000×g for 10 minutes, the supernatant was prepared as a protein extract. Equal amount of proteins were fractionated by electrophoresis on 8% or 10% acrylamide gels and were transferred onto a polyvinylidene fluoride (PVDF; Millipore, Billerica, USA) membrane followed by blotting with antibodies against Angptl4 (Sigma-Aldrich, St. Louis, USA), or β-actin (Sigma-Aldrich, St. Louis, USA). Protein levels were determined by using Western Breeze reagents (Santa Cruz Biotechnology, Dallas, USA) and Image Reader (LAS-3000 Imaging System, Fuji Photo Film, Tokyo, Japan). The secreted Angptl4 protein in the cell culture media were evaluated by enzyme-linked immunosorbent assay kits (R&D Systems, Minneapolis, USA).
Plasmid deoxyribonucleic acid and micro ribonucleic acid-132 transfection
Angiopoietin-like 4 plasmid DNA was purchased (Korea Human Gene Bank, Medical Genomics Research Center, KRIBB, Daejeon, Korea). BMCs were transfected with plasmid DNA or gWIZ mammalian expression vector (Genlantis, San Diego, USA) by using Lipofectamine 2000 (Invitrogen, Carlsbad, USA) according to the manufacturer's instruction. Micro ribonucleic acid-132 mimic and control miR, purchased (Bioneer, Daejeon, Korea), were transfected into hBMC using RNAiMAX transfection reagent (Invitrogen, Carlsbad, USA) according to manufacturer's protocol.
Statistical analysis
All data are presented as means±SDs. P were calculated by using the unpaired Student's t-test. For analysis of the in vivo ischemia experiments, the Scheffe's test was performed for multiple comparisons after ANOVA between the groups. A p<0.05 was considered statistically significant.
Results
Tube formation were reduced in DM-BMCs and recovered by oxytocin pretreatment
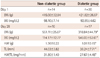
Hyperglycemia was induced 4 weeks after streptozotocin injection (Table 1), and the mortality was 32% in the diabetic group. Fasting blood glucose was substantially increased at day 28 (114.17±12.77 g/dL in non-diabetic group vs. 546.03±53.61 g/dL in diabetic group, p<0.01). The body weight was significantly reduced (551.11±25.87 g in non-diabetic group vs. 318.84±44.79 g in diabetic group, p<0.01) and the heart weight/tibia length ratio was significantly reduced in diabetic group (31.80±1.40 in non-diabetic group vs. 27.67±4.45 in diabetic group, p<0.01) (Table 1).
Angiopoietin-like 4, one of the diabetes-related genes, were involved in high glucose-insulted tube formation in BMCs
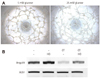
In a cDNA array, several genes were altered in their expression levels in DM-BMCs. Among them, we selected 9 genes which were rebounded to basal level in part (Table 2). After repeated reverse transcriptase-polymerase chain reaction, the expression patterns of 7 genes were analyzed and expressed as a graph. By the standard of reference line, Angptl4 and collagen, type 14, α1 (Col14α1) were upregulated, while tumor growth factor-β1 (Tgfb1), endothelial protein C receptor (Procr), endothelial cell-specific molecule 1 (Esm1), endothelin receptor type A (Ednra), and bone morphogenetic protein-binding endothelial regulator (Bmper) were downregulated (Fig. 2A).
The upregulated mRNA of Angptl4 was reduced in oxytocin-treated DM-BMCs (Fig. 2B).
High glucose insulted the human bone marrow-stem cells via angiopoietin-like 4 upregulation
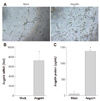
To perform the mechanistic study, the immortalized hBMCs were utilized. The tube formation was reduced in hBMCs with high glucose (25 mM) treatment (Fig. 3A). Angptl4 mRNA level was increased in high glucose-treated hBMCs for 9 days, and reduced in part by oxytocin treatment in high-glucose treated hBMCs (Fig. 3B).
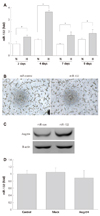
To examine the involvement of Angptl4 in hBMCs tube formation, Angptl4 was overexpressed in hBMCs. After two days of transfection, tube formation assay was performed. Few tubes were formed in Angptl4 expressed hBMCs in comparison with control cells (Fig. 4A). The mRNA of Angptl4 and protein of Angptl4 were increased by transfection with Angptl4 to hBMCs (Fig. 4B, C).
Micro ribonucleic acid-132 was involved in impaired tube formation via angiopoietin-like 4 upregulation
Endogenous miR-132 level was examined in hBMC with high glucose treatment over time. In comparison with normal glucose treated hBMCs, high glucose treated hBMCs increased miR-132 level. The level of miR-132 peaked at day 4, and returned to basal level (Fig. 5A). To examine whether miR-132 was directly related with angiogenetic activity in hBMCs, miR-132 was transfected for 2 days. miR-132 transfected hBMCs showed poor tube formation compared to miR control transfected hBMCs (Fig. 5B), and showed increased protein level of Angptl4 (Fig. 5C). Next, miR-132 level was measured after transfection with Angptl4 into hBMCs to determine if Angptl4 was the upregulator of miR-132. Angptl4-overexpressed hBMCs did not show the significant changes of miR-132 levels (Fig. 5D).
Discussion
Cell therapy with autologous BMCs is clinically feasible without ethical issues or immunological problems. Senescence or systemic conditions affect the application of autologous BMCs, and may contribute to the functional decline of stem cells. Several studies reported that diabetes impairs the neovascularization of BMCs.2)11)12)
In previous studies, we provided the evidence for beneficial roles of oxytocin in recovering the angiogenic potential via Kruppe-like factor-2 in DM-BMCs.2) To understand the mechanism that regulates stem cell function in the high glucose stress, we next explored which factor was contributing in disturbing the angiogenic potential. In microarray analysis, there were a number of genes which were altered in DM-BMCs and restored by oxytocin treatment. Our candidate genes were Angptl4, Tgfb1, Col14α1, Procr, Esm1, Ednra, Bmper, myogenic differentiation 1 (Myod1), protocadherin β5 (Pcdhb5), and secreted frizzled-related protein 4 (Sfrp4) (Table 2). Among them, we selected Angptl4 as a putative responsible gene.
Angiopoietin-like 4 is a secreted adipokine and involved in lipoprotein metabolism. Angptl4 modulate the vascular integrity in tissue-specific manner, and the role of Angptl4 in angiogenesis and vascular permeability remains controversial.13)14)15)16)
In DM-BMCs, Angptl4 was dramatically increased compared with non-DM-BMCs (Fig. 2). In other words, DM-BMCs were exposed to diabetic niches for 4 weeks, and it was suggested that Angptl4 might be induced by high glucose stress. To confirm the relationship of Angptl4 with high glucose stress, hBMCs were treated with 25 mM glucose. Additional interesting find was an upregulation of Angptl4 in hBMC by high glucose treatment (Fig. 3). These results showed the assured association of Angptl4 with high glucose stress.
So far, the precise role of Angptl4 in stem cells with regard to angiogenesis is not well understood, and we have investigated the effect of Angptl4 on angiogenesis activity in hBMCs. Overexpression of Angptl4 in hBMCs almost blocked the tube formation in angiogenesis assay. With these findings, Angptl4 could be suggested as a mediator of high glucose-induced angiogenic dysfunction in BMCs.
The functional consequences of miR-132 in endothelial cells are dependent upon an expression level of p120RasGAP, a negative regulator of Ras. In endothelial cells of tumor tissues, a large amount of miR-132 with minimal amount of p120RasGAP was present. On the other hand, the endothelial cells in normal tissue had abundant p120RasGAP without miR-132.17) MiR-132 has been proposed as an angiogenic switch in the endothelial cells targeting p120RasGAP to activate Ras and result in neovascularization.17)18) Recently, miR-132 delivery to human umbilical vein endothelial cells was reported to enhance the microvessels formation.19) In vascular smooth muscle cells, miR-132 targets LRRFIP1 to repress neointimal hyperplasia in rat carotid artery injury model.20)
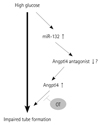
Micro ribonucleic Mir-132 is a novel angiogenic modulator in endothelial cells, and we investigated whether miR-132 was involved in high glucose-induced angiogenic dysfunction of hBMCs. The level of miR-132 was higher in high glucose-treated hBMCs than in normal glucose-treated cells. Unexpectedly, however, miR-132 transfected hBMCs showed no tube formation. Additional interesting find was that Angptl4 was increased by miR-132 transfection in hBMCs. In other words, Angptl4 was induced by both high glucose and miR-132 to exert negative effect on tube formation. To determine the sequence of the pathway, miR-132 was quantified in Agnptl4-overexpressed hBMCs, and there was no significant change in miR-132 levels. In summary, high glucose induced miR-132 induces Angptl4 via unknown Angptl4 antagonist to exert impaired tube formation in BMCs.
We presumed that the basal level of endogenous miR-132 would be a critical determinant of miR-132 for angiogenesis regulation. In this study, we did not compare the endogenous miR-132 levels of BMCs with other cell such as tumor cells and endothelial cells. To address the role of miR-132 on Angptl4 induction in BMCs, more systematic study is need.
Our data showed that the angiogenic function of BMCs might be disturbed by exposure to the diabetic microenvironment. In this study, we suggested that Angptl4 and miR-132 are involved in the significant aggravation of angiogenic function of high glucose-insulted stem cells (Fig. 6). Modulation of Angptl4 or miR-132 could be a promising option for functionally successful autologous cell therapy in diabetic patients.


 XML Download
XML Download