

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Kawasaki disease (KD) is a systemic vasculitis which develops exclusively in young children.1) Multiple genes are related to the pathogenesis of KD.2) The high incidence of KD in Asian ethnic groups3) has stimulated extensive searches for the genetic susceptibility for KD. Most of the susceptible genes are related to inflammation and aberrant immune reactions.4) Whatever the initiating agent of inflammatory reaction is, the individual differences of a series of immunologic inflammatory cascades due to a given genetic polymorphism are the primary focus of etiologic research on KD.5) Interleukin-6 (IL-6) gene polymorphisms,6)7) tumor necrosis factor-α (TNF-α) gene polymorphisms,8) CD40 ligand gene polymorphisms,9) genetic polymorphisms of encoding chemokine receptors,10) programmed death-1 (PD-1) gene polymorphisms,11) and other genome wide association studies have been researched.2)
In addition, the paucity of regulatory T cells (Treg cells)12)13) and the plasticity of T cell populations14) have been investigated in the development of KD. Recently T follicular helper cells (Tfhcells) have been a focus of interest in autoimmune vasculitis.15) Tfhcells are involved in the germinal center formation for effective B cell differentiation to antibody-producing plasma cells. T cells and B cells have an effect on each other in every step of adaptive immune reaction. Therefore, we concentrated on Tfhcells as an emerging factor for balancing T cell plasticity after infection15) and affecting B cell development. King et al.16) reported that homeostatic expansion of T cells during immune insufficiency generates autoimmunity. Therefore, repetitive viral infections or potent live vaccinations in infants may induce relative immune insufficiency and high turnover of T cells may direct to autoimmunity, the state of KD. Tfhcells work effectively via IL-21/IL-21 receptor (IL-21R) pathway.15) IL-21 promotes B cell differentiation but if there is a defect in IL-21R, IgE is increased and immunoglobulin class switching recombination is decreased.17) Jin et al.17) reported that IL-21 directs B cell growth arrest for nonspecifically or inappropriately activated B cells while normally it promotes B cell maturation during T cell dependent B cell activation. Therefore, in KD, we could infer that IL-21 may arrest growth of B cell through lacking of IL-21R expression due to IL-21R gene polymorphism. Ineffective B cell differentiation is prominent in KD compared with other febrile diseases.18) IL-21 may be increased due to ineffective B cell differenciation19) but lack of IL-21R may lead to functional defect in immunoglobulin switching and increased IgE level.
Now, we speculated that KD might develop in certain young individuals who have IL-21R gene polymorphism when IL-21R expression down regulated transiently, and immunoglobulin switching may be arrested by ineffective IL-21R expression. In advance we checked the serum level of IL-21 in patients with KD and it was increased.20) Hence, we investigated the IL-21R gene polymorphisms in the promoter region which could decrease the expression of IL-21R gene in aspect of the genetic susceptibility of KD.
Go to : 
Subjects and Methods
Subjects
A total of 200 samples (100 samples of patients with KD and 100 of healthy controls) were studied from February 2010 to October 2011 in Wonju Christian Hospital, Wonju, South Korea. KD was diagnosed according to KD criteria.21) The control group was composed of healthy adults who were recruited into a hypertension cohort study and had no medical history of KD or autoimmune disease and have normal blood pressure. Whole blood was collected after receiving informed consent. This study was approved by the Institutional Review Board of Hospital, Yonsei University (CR309043).
Deoxyribonucleic acid extraction, polymerase chain reaction, and sequencing
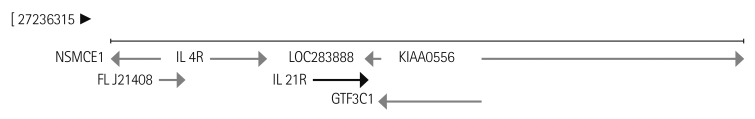
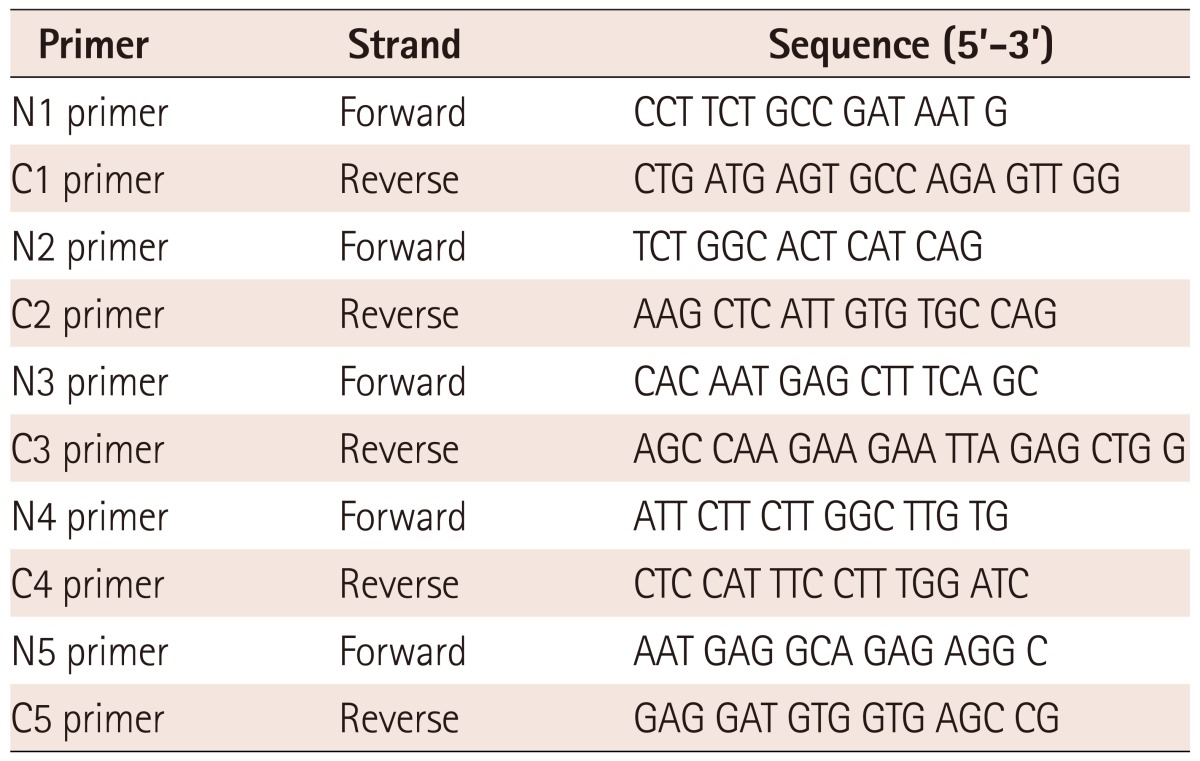
Genomic deoxyribonucleic acids were extracted from whole blood samples of patients and controls by using LaboPass™ Blood Kit (Cosmo Genetech, Seoul, Korea) according to the manufacturer's instruction. The nucleotide changes in the promoter region of IL-21R gene (-2500 bp~ +1 bp) according to the position from the transcription starting base of the IL-21R gene (NG_012222.1) (Fig. 1) were identified by PCR using ExTaq polymerase (Takara Bio Inc, Shiga, Japan) with 5 pairs of primers listed in Table 1. PCR conditions were 1 cycle of 3 minutes at 95℃, followed by 35 cycles of 3 steps that were at 95℃ for 30 seconds, at 58℃ for 30 seconds, at 72℃ for 1 minute each, and 1 cycle of 10 minutes at 72℃. The PCR products were separated by gel electrophoresis on 1% agarose gel.
We screened the known single nucleotide polymorphism (SNP) sites for IL-21R in patients with KD: rs11074858, rs56189459, and rs2214032 for IL-21R gene, which are located in exons. However there was no significant change of these genetic sites in patients with KD. Therefore we studied the promoter region of IL-21R. Among the 5 experiments for screening SNP of promoter genes, we found significant SNPs in -500 bp~ +1 bp region. We analyzed the sequences using direct sequencing method using ABI 3730 capillary sequencer (Applied Biosystems, CA, USA) and compared sequences with reference sequence through Human BLAT program (developed by Jim Kent in UCSC Genome Browser).
Measurement of interleukin-21 and total IgE in sera
We measured the serum IL-21 level in patients with KD by enzyme linked immunosorbent assay kits (e-Bioscience, San Diego, CA, USA) and total IgE by immunoassay system (Siemens, Munich, Germany).20)
Statistical analysis
Sample size was determined by the PASS program 2008 (NCSS Statistical Software, Kaysville, UT, USA). The difference of allele frequencies between KD patients and controls was compared by Fisher's exact test using GraphPad Prism version 5.00 for Windows (GraphPad Software, San Diego, CA, USA). The difference of serum level of IL-21 or total IgE according to the presence of IL-21R gene polymorphisms was calculated by Mann-Whitney test. The difference of prevalence of coronary arterial dilatation (CAD) according to IL-21R gene polymorphisms was calculated by Fisher's exact test. Statistical significance was defined as p<0.05.
Go to :
Results
Characteristics of patients with Kawasaki disease
A total of 100 patients and 100 healthy controls were enrolled in this study. The median age of patients with KD was 3 years of age (range: 0.3-11.0) and gender ratio (male : female) was 55 : 45 (1.2 : 1). The gender ratio of healthy controls was 1 : 1. The proportion of atypical KD patients was 28% and that of refractory KD patients was 6%. The number of patients with CAD when the echocardiogram was performed in febrile period was 29 (29%).
Single nucleotide polymorphisms in the promoter region of interleukin-21 receptor gene and nomenclature
Among the SNPs in the promoter region of IL-21R gene, SNPs of which minor allele frequency (MAF) >0.01 were depicted in Table 2. Most SNPs were located in -500 bp~ +1 bp from transcription starting site. In this study, we could not find the known SNPs of this region such as rs2189522 or -83 T>C in both groups. Several SNPs which had not been reported in dbSNP site were found. Therefore, we named these SNPs as either the chromosome position in the chromosome 16 region (NC_000016.9) or the position from the transcription starting base of the IL-21R gene (NG_012222.1).
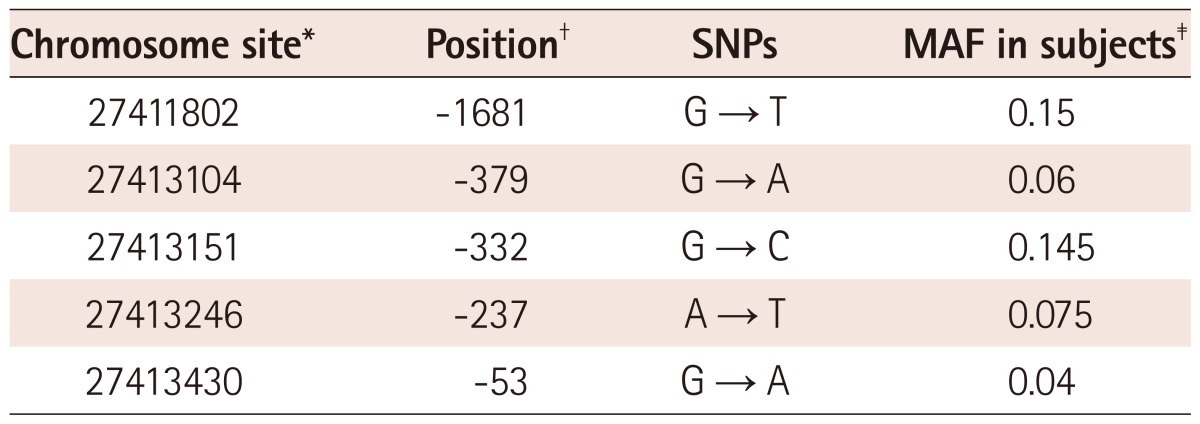
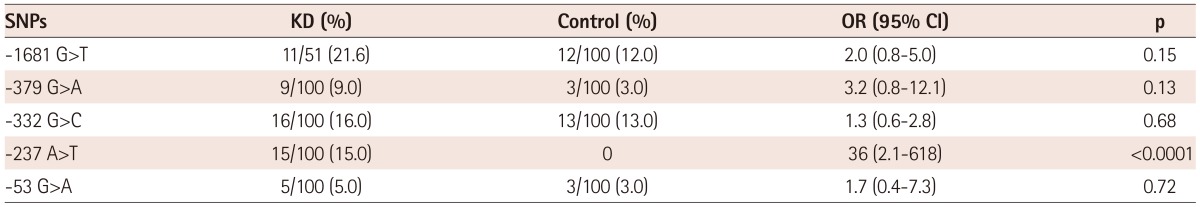
Differences in allele frequencies of single nucleotide polymorphisms between patients with Kawasaki disease and controls
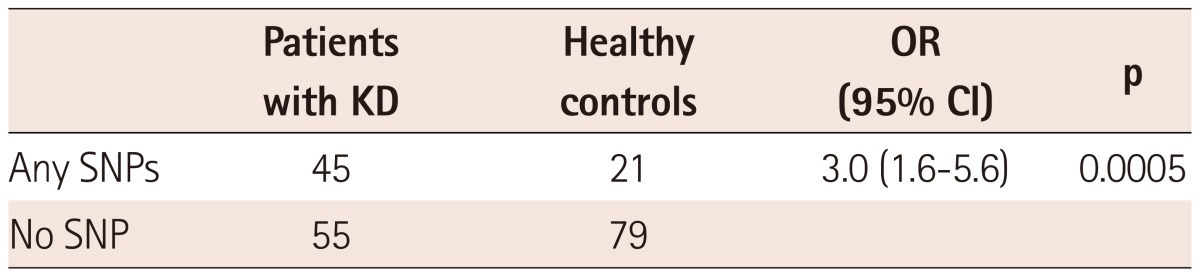
Table 3 shows the allele frequencies of SNPs in patients and control groups. Among the SNPs, -237 A>T polymorphism was found in 15 patients with KD only and significant difference in allele frequency between the patients and the controls was found {OR: 36, 95% confidence interval (CI): 2.1-618, p<0.0001}. Because there were more SNPs of which MAF <0.01 and one individual had more than one SNP, we divided the study population into individuals with any SNPs and those with no SNP (Table 4). There was a significant difference in terms of number of individuals who have any SNPs of IL-21R gene between the patients group and controls (OR: 3.0, 95% CI: 1.6-5.6, p=0.0005).
Relationships between interleukin-21 receptor gene polymorphisms and the serum levels of interleukin-21
When we searched for differences in the serum level of IL-21 by the presence of IL-21R gene polymorphisms in 25 patients with KD, no significant difference was shown (Fig. 2). The mean serum level of IL-21 in patients who have polymorphisms was 612±480 pg/mL and that of patients who have no polymorphism was 414±290 pg/mL, there was no significant difference between two groups (p=0.18).
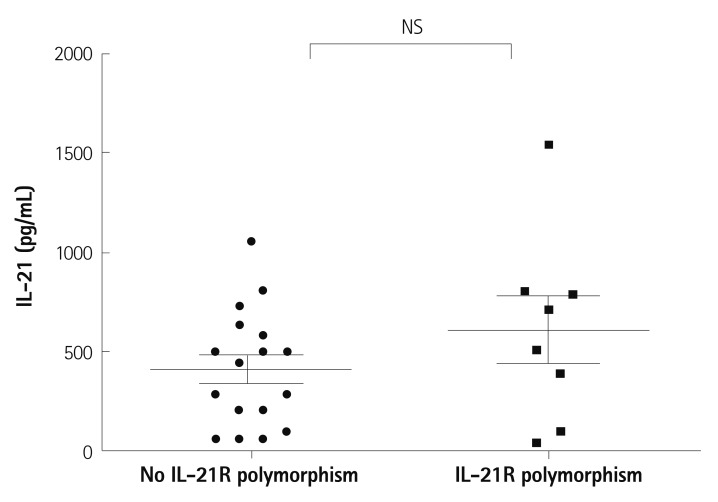 | Fig. 2Difference of serum levels of IL-21 according to the presence of single nucleotide polymorphisms of IL-21R gene. The mean serum level of patients who have polymorphisms was 612±480 pg/mL and that of patients who have no polymorphism was 414±290 pg/mL, there was no significant difference between two groups (p=0.18). NS: not significant, IL-21: interleukin-21, IL-21R: interleukin-21 receptor.
|
Association of interleukin-21 receptor gene polymorphisms and total IgE
Serum levels of total IgE were also checked in the same group above. There was no significant difference in terms of total IgE level between patients with polymorphisms and those with no polymorphism (Fig. 3). Our data couldn't show the significant difference of the median values of serum total IgE between the patients who have IL-21R gene polymorphisms and those who have not (30.1 pg/mL vs. 58.6 pg/mL, p=0.17).
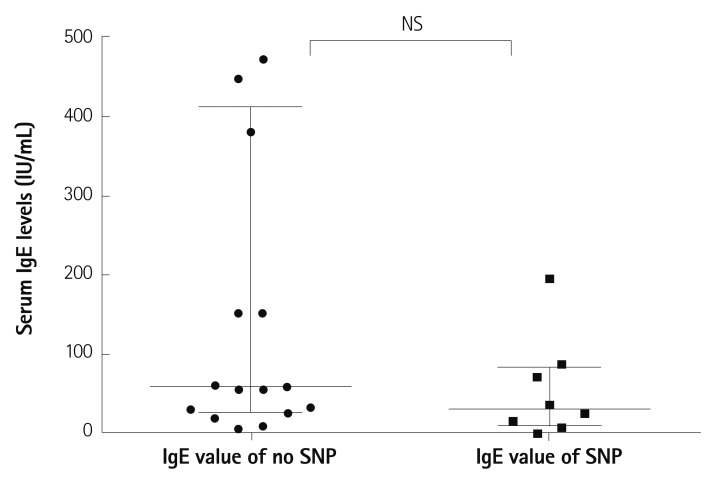 | Fig. 3Serum levels of total IgE according to IL-21R gene polymorphisms in patients with KD. Our data showed no significant difference of serum total IgE between the patients who have IL-21R gene polymorphisms and those who have not (median value=30.1 pg/mL vs. 58.6 pg/mL, p=0.17). NS: not significant, IgE: immunoglobulin E, IL-21R: interleukin-21 receptor, KD: Kawasaki disease.
|
Interleukin-21 receptor gene polymorphisms and coronary arterial dilatation
The prevalence of CAD in the group who had IL-21R gene polymorphisms and that had not IL-21R gene polymorphisms were not significantly different (OR=0.99, 95%CI=0.42-2.4).
Go to :
Discussion
The IL-21/IL-21R pathway is known as an executor of B cell differentiation.22) IL-21 is produced when the immune reaction reaches its peak by CD4+ T cells and IL-21R is expressed in mature B cells. IL-21 induces PD-1 expression on T cells and plays a role in making a knot of a certain immunologic cascade. To our knowledge, this is the first study to investigate IL-21R gene polymorphisms in KD. We found significant gene polymorphisms in patients with KD compared with controls. Because IL-21 is increased by autocrine manners,23) we assumed that the structure of IL-21R would be still intact even in the presence of polymorphism. In the preliminary study, we couldn't find any significant polymorphisms belonging to 9 exons of IL-21R gene in patients with KD. Therefore we searched the promoter region of IL-21R gene. Pène et al.22) studied -83 T>C in atopic patients and controls and they found more frequent variants in atopic patients compared with controls. Their functional study showed that IL-21R gene polymorphism may increase the serum level of total IgE against the IFN-γ mediated inhibition of IgE synthesis. In our study, 30% of patients with KD had any allergic history and family history of any allergic diseases (data not shown). Our data showed no correlation between IL-21R gene polymorphism and the serum level of total IgE or serum level of IL-21. This result may be due to many other molecules which could affect the serum level of IL-21 such as PD-1, SOCS-1, SAP, Blimp-1, Bcl-6 and WSB-2.24-28) Therefore, elevated serum level of IL-21 may not be described only by IL-21R gene polymorphisms. However, our data contribute to the understanding of one of the aberrant immune reactions in KD. No allergy, cytotoxic T cell mediated inflammation or inappropriately activated macrophages can entirely explain the inflammatory reactions of KD. We hypothesized that the polymorphisms of IL-21R gene may be responsible for decreased expression of IL-21R. In promoter region of IL-21R gene, 27413104 G → A change makes the sequence as GAAC/CAAG a symmetric mirror sequences. We don't know what this change means, but this change may be responsible for the altered the transcription of IL-21R gene. Further functional study should be required to prove the function of SNPs that we found.
There are patients who had more than 2 SNPs. Compared with healthy controls, patients with KD have more combinations of SNPs in IL-21R gene. Although there is no significant difference of MAF of each SNPs between patients group and healthy controls, the frequency of combination of SNPs was higher in patients group than in controls.
We could not find a significant increase of known SNPs of IL-21R gene29) in this study. Those SNPs had been studied in Caucasian ethnic groups, therefore our results may be explained by ethnic differences.
We have no preventive strategy for KD yet. However, we might speculate that repeated infections or vaccinations may down-regulate the IL-21R and this effect would be aggravated in hosts who have IL-21R gene polymorphisms, which may lead to arrest of B cell differentiation and elevated serum levels of total IgE and IL-21. Therefore, avoiding frequent infections in a short term period may be a feasible strategy for preventing KD.
In addition to this, we cautiously suggest that anti-IL-21 antibody could be a new therapeutic agent for IVIG-resistant KD in patients who have IL-21R gene polymorphisms. Anti-IL-21 antibody is being studied in autoimmune diseases as a new therapeutic agent.30)
In conclusion, our data suggest that the genetic susceptibility profile for KD may include IL-21R gene.
Go to :
 XML Download
XML Download