

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Restenosis after percutaneous intervention for coronary artery disease is due to the intimal tissue growth caused by the migration and proliferation of vascular smooth muscle cells (VSMCs).1)2) Brachytherapy3)4) and drug-eluting stents5)6) have been introduced to prevent this proliferation.
Vascular endothelial growth factor (VEGF) is a potent endothelial cell-specific mitogen, which enhances vascular permeability and stimulates angiogenesis.7)8) Therapeutic angiogenesis, using VEGF gene transfer, has been performed in selected patients with critical limb ischemia.9)10) Studies have also been performed with VEGF for ischemic heart disease.11-13) VEGF direct gene transfer may passivate endovascular stents by accelerating stent endothelialization. thereby reducing in-stent thrombus formation and obstruction due to intimal thickening.14)15) However, Swanson et al.16) reported that VEGF-eluting stents did not accelerate re-endothelialization or inhibit restenosis.
Recent experimental studies have demonstrated the collaterogenic effect of monocytes/macrophages, T lymphocytes, bone marrow-derived progenitor cells, VEGF family members, tumor necrosis factor-alpha and basic fibroblast growth factor. However, the Janus phenomenon implies that these cells and molecules may worsen atherosclerosis.17) VEGF may have an atheroprotective effect because it protects endothelial cells against the toxic effects of oxidized low density lipoprotein; however, by securing endothelial integrity and survival, VEGF may conversely favor plaque growth and destabilization.18)
VEGF regulates angiogenesis by primarily interacting with main two tyrosine kinase receptors, VEGF receptor-1 (VEFGR-1, also known as Fms-like tyrosine kinase, Flt1) and VEGF receptor-2 (VEGFR-2, also known as fetal liver kinase, Flk1).19) VEGFR-1 is expressed on vascular endothelial cells as well as non-endothelial cells, including smooth muscle cells, monocytes, trophoblasts, mesangial cells and osteoblasts. VEGFR-2, which has tyrosine kinase activity, is mostly located on vascular endothelial cells and is an important factor under both physiologic (wound healing, ovulation and menstruation) and pathologic conditions (tumor growth).18)
By screening peptide libraries, a soluble arginine-rich hexapeptide (e.g., RRKRRR, dRK6) and anti-flt-1 (Gly-Asn-Gln-Trp-Phe-Ile or GNQWFI) were synthesized at Pohang University of Science and Technology, Korea.20)21) These workers reported that dRK6 can bind VEGF and; thereby, block the interaction between VEGF165 and VEGF receptors as well as the growth and metastasis of VEGF-secreting HM7 human colon carcinoma cells in nude mice.20) Moreover, anti-flt-1 peptide has shown specificity for VEGFR-1 binding and inhibitions of VEGF, placental growth factor (PlGF) and VEGF/PlGF heterodimer binding to VEGFR-1.21)
Shibata et al.22) have shown that VEGF is involved in the process of restenosis, due to its angiogenic properties, and induction of myocyte chemotaxis in pigs following stenting. However, it is not clearly known whether the above-mentioned anti-VEGF peptides, dRK6 and anti-flt-1, suppress neointimal formation following percutaneous intervention. Therefore, whether the neointima formation is inhibited by dRK6 (a VEGF antipeptide) or anti-flt-1 (a VEGFR-1 antipeptide) in balloon-injured rat carotid arteries was investigated.
Materials and Methods
Reagents
The dRK6 and anti-flt-1 were synthesized at Pohang University of Science and Technology, Korea, as described previously.20)
Animals
Male Sprague-Dawley rats, weighing 300-350 g, were used in this experiment, and were subcutaneously injected with 0.5 mg/kg of dRK6 (N=5) or 0.5 mg/kg of anti-flt-1 (N=5), which were dissolved in phosphate buffer solution (PBS), 2 days prior to induction of a carotid balloon-injury, and then daily for 2 weeks post carotid balloon injury. Control animals received PBS alone (N=5) or the reverse peptide of dRK6 (N=4), again daily for 2 weeks post carotid injury.
Carotid balloon-injury model
Arterial injury was induced using the balloon denudation technique.23) After induction of anesthesia with an injection of xylazine (4.6 mg/kg) and ketamine (70 mg/kg), a cervical midline incision was made, with the bifurcation of the left carotid artery then exposed. Blood flow to the site of the surgical manipulation was temporarily interrupted by lifting the left common, internal and external carotid arteries with the use of surgical ligatures. A 2 Fr balloon catheter (Fogarty, Baxter, USA) was then introduced via an arteriotomy in the external carotid artery and advanced to the proximal edge of the omohyoid muscle. To produce denudation and a mechanical stretching injury of the left common carotid artery, the balloon was inflated to generate slight resistance, and then withdrawn from the proximal part of the omohyoid to the carotid bifurcation three times. Following this procedure, the catheter was removed and the insertion ligated.
Tissue preparation
Two weeks after induction of the balloon injury, animals were euthanized, and the left carotid and the left femoral arteries exposed. Via a puncture in the left femoral artery, a catheter was introduced into the ascending aorta, with the aorta fixed with 4% formaldehyde at 90-100 mmHg for 5 minutes. Following the initiation of perfusion and fixation, animals were euthanized with an overdose of thiopental sodium via a tail vein. Left carotid arteries were stored in 4% formaldehyde for 24 hours, and then embedded in paraffin for sectioning at thicknesses of 5 µm. Hematoxylin-eosin staining, elastin fiber staining and immunochemical studies were then conducted.
Histomorphometric study
After recording digital images at ×100 magnification, the luminal (the area circumscribed by the intimal border), intimal (the area between the lumen and the internal elastic lamina) and the medial (the area between the internal and external elastic lamina) areas were delineated by computerized morphometry (AxioVsion LE v 4.1.1, Carl Zeiss Vision GmbH, Germany).
Immunohistochemistry
To detect proliferative cells, bromodeoxyuridine (BrdU) (Sigma-Aldrich Co, St. Louis, MO, USA) was injected intramuscularly 2 hours (100 mg/kg) before euthanasia. Immunohistochemistry was performed using a monoclonal antibody (Dako Cytomation California Inc., Carpinteria, CA, USA) against BrdU. BrdU-labeled cells in the intima were counted at ×400 magnification. BrdU-labeling indices (the fraction of labeled nuclei times 100) were calculated in the intima to assess the rate of cell replication. In addition, immunohistochemistry was performed using antibodies against VEGF (A-20: sc152, Santa Cruz Biotechnology, Santa Cruz, CA, USA), VEGFR-1 (Flt-1, Lab Vision, Fremont, CA, USA), VEGFR2 (Flk-1/DR/VEGFR2, Lab Vision), monocyte chemoattractant protein-1 (MCP-1, anti-mouse CCL2/MCP-1 antibody, R&D Systems, Minneapolis, MN, USA) and IL-6 (anti-rat IL-6 antibody, R&D Systems). Reaction products were developed with diaminobenzidine (Sigma-Aldrich Co) and counterstained with hematoxylin.
Results
Inhibitory effects of dRK6 or anti-flt-1 peptide on neointimal formation
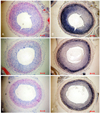
The carotid arteries in the control group developed significant neointimal formation by day 14 (Fig. 1), where those in the dRK6 and anti-flt-1 groups showed less neointimal formation (neointimal area; control group 0.44±0.09 mm2, dRK6 group 0.25±0.05 mm2 and anti-flt-1 group 0.19±0.05 mm2, p<0.05) (Fig. 1, 2) (Table 1).
Increased expressions of vascular endothelial growth factor and vascular endothelial growth factor receptor-1 in injured carotid arteries
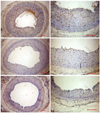
The localizations of VEGF and VEGFR-1 were immunohistochemically studied in rats. VEGF and VEGFR-1 were immunopositive in the nuclei of neointimal cells in the control group on day 14 (Fig. 4). The administration of the anti-flt-1 peptide showed a slight effect on the expression of VEGF, and both dRK6 and anti-flt-1 reduced the VEGFR-1 immunoreactivities. VEGFR-2 was expressed on the luminal side of the control group, but was absent in the dRK6 and anti-flt-1 groups (Fig. 4).
Inhibitory effects of dRK6 or anti-flt-1 peptide on inflammatory changes
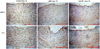
Immunohistochemical staining performed 14 days after balloon injury showed increased monocyte chemoattractant protein-1 (MCP-1) and IL-6 immunoreactivities in neointimal cells, which were attenuated by dRK6 and anti-flt-1 (Fig. 5).
Discussion
In contrast to VEGF and its receptor, VEGFR-2, PlGF and its receptor, VEGFR-1, have been poorly investigated and largely neglected.18) However, recent gene-targeting studies have indicated that PlGF and VEGFR-1 are key regulators of the angiogenic switch under pathological conditions.18)24) The anti-VEGFR-1monoclonal antibody (mAb) has been reported to block neovascularization in the ischemic retina18)25) and to dose-dependently block angiogenesis and growth of human epidermoid A431 tumors in nude mice.26) Autiero et al.18) used anti-VEGFR-1 and anti-VEGFR-2 mAb to study their effects on the growth and stability of initial (avascular) fatty streak lesions as well as interme-diate and advanced (vascularized) plaques in atherosclero-sis-prone apolipoproteinE-deficient mice. Treatment with anti-VEGFR-1 mAb was found to reduce the sizes of the early and intermediate lesions as well as the growth of advanced atherosclerotic lesions. However, anti-VE-GFR-2 mAb failed to affect atherosclerotic plaque deve-lopment at any stage. However, surprisingly, neither anti-VEGFR-1 mAb nor anti-VEGFR-2 mAb blocked angiogenesis in atherosclerotic lesions or the surrounding adventitia. Therefore, it appears anti-VEGFR-1 mAb suppressed plaque growth and vulnerability by inhibiting inflammatory cell infiltration, independent of angiogenesis; whereas, anti-VEGFR-2 mAb, which normally blocks angiogenesis, was ineffective. It is also important to note that the inhibition of inflammatory cell infiltration is related to the suppression of plaque growth.
In this study, anti-peptides, instead of monoclonal antibodies, were used to block either VEGF or VEGFR-1 in the balloon-injured rat carotid artery. Yoo et al.27) demonstrated that dRK6 suppressed ongoing paw inflammation in collagen-induced arthritis and blocked the VEGF-induced production of proinflammatory cytokines in a mouse model. Moreover, the anti-flt-1 peptide (Gly-Asn-Gln-Trp-Phe-Ile or GNQWFI), which inhibits VEGFR-1 binding, was also identified by screening a synthetic peptide library.21) The anti-flt-1 peptide was found to bind specifically with VEGFR-1, but inhibit the bindings of VEGF, PlGF and the VEGF/PlGF heterodimer to VEGFR-1. Moreover, anti-flt-1 peptide effectively blocked the VEGF-induced migration of endothelial cells and their ability to form capillarylike structures on a fibrin gel-based in vitro angiogenesis system.21) Furthermore, the growth and metastasis of VEGF-secreting tumor cells were also significantly inhibited by subcutaneous injections of anti-flt-1 peptide in nude mice.21) The blocking of either VEGF or VEGFR-1 by anti-peptides has some advantages over gene transfer or the use of a monoclonal antibody; it does not use a viral vector, does not cause an antigen-antibody reaction and is less expensive. The present study has demonstrated that both the blocking of VEGF by dRK6 and VEGFR-1 by anti-flt-1 peptide reduces neointimal formation in a balloon-injured rat artery. To our knowledge, this is the first study to evaluate the effect of dRK6 or anti-flt-1 peptide on neointima formation following induction of a balloon-injury.
Ohtani et al.28) reported that the increased expression and activity of VEGF are essential for the development of experimental restenosis following an intraluminal injury, due to the recruitment of monocyte-lineage cells; and that blockade of VEGF by soluble VEGF receptor 1 gene transfer attenuated neointimal formation following an intraluminal injury and inhibited the increased expression of inflammatory factors, such as monocyte chemoattractant protein-1 (MCP-1) and VEGF in rabbits, rats and mice. Yamada et al.29) showed that VEGF-mediated angiogenesis and inflammation are actually mediated by monocyte chemoattractant protein-1 (MCP-1). This present study also showed that MCP-1 was reduced in the dRK6 and anti-flt-1 peptide groups, which supports the role of MCP-1 in neointimal formation following vascular injury and the suppression of neointimal formation due to dRK6 and anti-flt-1 peptide.
De Leon et al.30) reported a BrDU labeling index of 27.5±3.8% in balloon injured rat carotid arteries on day 3 post-injury. In the present study, BrDU positive cells in the neointima were reduced in the dRK6 and anti-flt-1 peptide groups compared to the control group on day 14 post-injury, suggesting VEGF anti-peptides suppressed neointimal proliferating cells post-injury.
There was no difference between anti-flt-1 peptide and dRK6 with respect to reduced neointimal formation, but this will require further elucidation. The re-endothelialization in a rat carotid artery following a balloon injury was not conducted in this study, but this might also be affected by VEGF inhibition. Swanson et al.16) reported that a VEGF-eluting stent did not accelerate re-endothelialization or inhibit restenosis, but appeared to reduce stent thrombosis. Theoretically, inhibition of VEGFR-1 by anti-flt-1 peptide might be more beneficial than the blocking of VEGF itself by dRK6, as this does not affect VEGFR-2 mediated endothelialization. In the present study, the dRK6 group tended to show an increased incidence of thrombosis (data not shown), which may have been due to the suppression of re-endothelialization by dRK6. Studies on the side effects of anti-flt-1 or dRK6 on the possible thrombogenic potential, as well as their effects on angiogenesis and re-endothelialization are still required. Also, the immunoreactivities of MCP-1 and IL-6 were not quantitatively analyzed in this study.
Many studies have been performed on the effects of blocking VEGF or VEGFR-1, using anti-peptides, in cancer and rheumatic disease, but no study has been conducted on the use of anti-peptides to reduce restenosis. Therefore, anti-VEGF peptides were concluded to potentially offer a route for the development of therapeutic agents that will prevent restenosis following percutaneous coronary intervention.



 XML Download
XML Download