

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Platelets, the fundamental component of primary hemostasis, are also known to be reservoirs for many growth factors (GFs), which they store in their α-granules. Platelet aggregation and activation, after vascular damage, result in the release of several GFs that may affect the chemotaxis, proliferation, and differentiation of mesenchymal stem cells (MSCs) or other committed cells during the process of tissue repair and healing [1]. The GFs released from platelets include platelet-derived growth factors (PDGFs), transforming growth factor-beta (TGF-β), vascular endothelial growth factor (VEGF), epidermal growth factor (EGF), fibroblast growth factor (FGF) and insulin-like growth factors (IGFs) [2, 3].
To explore the possibility that platelet-rich plasma (PRP) could provide an autologous source of these essential GFs that benefit bone and soft tissue healing [4], many clinical and experimental studies dealing with the effects of PRP have been conducted [5-7]. However, the benefit of PRP on bone formation is a controversial subject. While a report suggested a stimulatory effect with the addition of PRP [5], others have observed no improvement [6] or have detected even inhibitory effects [7]. Although the lack of standardization in application across these studies, including differences in the preparation method or dosage of PRP, biomaterials, species, implantation sites, and cell types, may have contributed to the inconsistent results [8], we hypothesized that the differences in GF composition among PRPs could lead to this discrepancy. GF quantities vary significantly between individuals [9]. The potential efficacy of PRP depen-ds on the levels of GFs released from platelets; however, no study has yet addressed the relationship between the biologic effects and GF concentrations in PRP. In this study, we attempted to explore the variations in GF release patterns in PRP in order to determine whether differences in GF concentrations influence biologic effects such as proliferation and alkaline phosphatase (ALP) activity of human MSCs (HMSCs) and to identify the factors associated with each of these biologic effects.
Go to : 
MATERIALS AND METHODS
This study was reviewed and approved by the Institutional Review Board of Yeungnam University College of Medicine. Prior to drawing blood or bone marrow, informed consent was obtained from each donor.
1. Preparation of PRP and measurement of GF concentrations
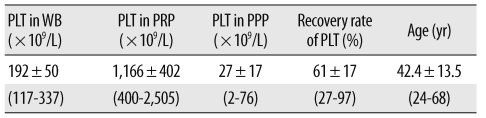
First, PRPs were obtained from 3 healthy male volunteers (27 yr, 39 yr, and 43 yr old) to explore the effects of PRP concentration on the proliferation and ALP activity of HMSCs. Then, we obtained donated PRPs from 39 other healthy volunteers to assess the relationship between the effects and GF concentrations. The mean donor age was 42 yr (range, 24-68 yr), distributed at 6-8 individuals per decade. We limited our subjects to those who had not taken anti-platelet medications within 1 week of donation; further, to minimize the possible influences of hormonal variation on the proliferation of HMSCs, we included only male donors.
PRPs were prepared via double centrifugation of blood. In brief, 9 mL of venous blood was drawn into a polypropylene tube containing 1 mL of anticoagulant, acid citrate dextrose (ACD). The blood was centrifuged at 1,500 rpm for 10 min at 25℃ to separate the platelet-containing plasma from the red cells. The platelet-containing plasma was then centrifuged at 3,000 rpm for an additional 10 min at 25℃, and platelet-poor plasma (PPP) was separated out. Platelet pellets were resuspended in 1 mL of plasma and were pooled for PRP. To activate the platelets, 1 part bovine thrombin stock solution (1,000 U/mL; Sigma, St. Louis, MO, USA) was added to 9 parts PRP or PPP to yield a final thrombin concentration of 100 U/mL. Each sample was incubated for 1 hr at 37℃ [10]. The resulting supernatants from the clot preparation were referred to as activated PRP (aPRP) and activated PPP (aPPP). Both aPRPs and aPPPs were stored at -80℃ until use. The concentrations of PDGF-AA, PDGF-AB, PDGF-BB, TGF-β1, FGF-basic (FGF-b), IGF-1, and VEGF were measured with commercially available Quantikine colorimetric sandwich ELISA kits (R&D Systems, Minneapolis, MN, USA) according to the manufacturer's instructions.
2. Cell isolation
HMSCs were cultured using established protocols [11]. Bone marrow aspirate was obtained with consent from a 50-yr-old female patient who had undergone an autologous bone graft for the treatment of degenerative spondylolithesis. Mononuclear cells were isolated using a density gradient and were cultured in DMEM (Sigma, St Louis, MO, USA) with 10% fetal bovine serum (FBS), penicillin, and streptomycin (Gibco-BRL, Rockville, MD, USA). The cells were incubated in a humidified atmosphere of 5% CO2 at 37℃. Non-adherent cells were discarded after 24 hr, and the medium was exchanged every 3 days. First passage (P1) cells were utilized in the study.
3. The effects of aPRP and aPPP on the DNA content and ALP activity of HMSCs
The cells were seeded at a density of 1.5×104 cells/well in 12-well plates (35 mm diameter) and were allowed to attach for 24 hr. After this adhesion period, the medium was removed and was replaced with DMEM with 10% FBS (as control) or variable volume concentrations of aPRP and aPPP (1%, 3%, 10%, and 30%) from each of the 3 donors. Each experiment was conducted in triplicate. Media was exchanged every 4 days. The level of DNA and ALP activity were measured in quadruplicate on the fourth and twelfth days. The DNA level was measured via Hoechst DNA stain, and ALP activity was determined by quantifying the p-nitrophenol released from p-nitrophenol phosphate [12].
4. Relationships between GF concentrations, DNA content, and ALP activity of HMSCs
In order to evaluate the effects of aPRP GF concentrations on the proliferation or ALP activity of HMSCs, the HMSCs were cultured under the same conditions as described above, in DMEM with 10% aPRPs from each of the 39 donors. After culturing for 12 days, the DNA contents and ALP activity were measured.
5. Statistical analysis
Statistical analysis was conducted using the Predict Analytics Software program (PASW) version 18.0 (SPSS Inc., Chicago, IL, USA). The Shapiro-Wilk test was used to analyze the normal distribution of the data. The effects of aPRP and aPPP concentrations on the DNA content and ALP activity of HMSCs were evaluated via ANOVA, or the Kruskal-Wallis (K-W) test with Bonferroni correction. The differences in GF concentrations between aPRPs and aPPPs were examined using the paired t-test. Pearson's correlation analysis was used to identify correlations between GF levels and age, platelet count, or mean platelet volume. The aPRP release pattern of GFs was grouped via cluster analysis using the K-means method. The relationships between GF concentrations and the biologic effects between these groups was analyzed using ANOVA or K-W test with Bonferroni correction. Stepwise multiple linear regression analysis was employed to show which GFs were associated with DNA content or ALP activity. Significant differences were established at P<0.05; the Bonferroni corrected P value threshold (0.05/number of comparison) was employed for post-hoc analysis of the K-W test.
Go to :
RESULTS
1. Effects of PRP and PPP on the DNA content and ALP activity of HMSCs
On the fourth day after cultivation, DNA contents were unaffected by the addition of aPRP (P=0.043 by K-W test, but multiple comparisons with Bonferroni correction did not reveal significant differences). The addition of aPPP did not affect DNA content at any concentration (P=0.408 by ANOVA). However, by the twelfth day, DNA levels differed significantly according to the aPRP and aPPP concentrations. Both aPRP and aPPP, at a concentration of 10% resulted in increased DNA levels. However, DNA levels were reduced by aPPP at a concentration of 30% (P<0.001 by ANOVA, post-hoc analysis using Scheffe method; Fig. 1A). The stimulatory effects of aPRP were significantly greater than those of aPPP (P=0.006 by paired t-test). The ALP activity of cultured HMSCs was reduced with the addition of aPRP (P<0.001 by ANOVA), but the addition of aPPP did not affect the cellular ALP activity (P=0.564 for the fourth day; P=0.094 for the twelfth day; both by ANOVA; Fig. 1B).
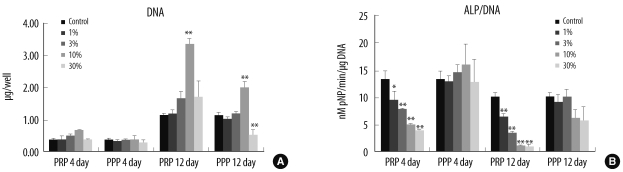 | Fig. 1The DNA concentration (A) and ALP activity (B) of human mesenchymal stem cells after the addition of 1%, 3%, 10%, and 30% activated PRP or activated PPP. Each value is the mean of 3 donors, with the error bar showing one standard deviation (*P<0.05; **P<0.01 compared with control).
Abbreviations: ALP, alkaline phosphatase; PRP, platelet-rich plasma; PPP, platelet-poor plasma.
|
2. Characteristics and concentrations of GFs in aPRP and aPPP
Plasma samples from 39 individuals were evaluated in this portion of the study. Platelet counts in native whole blood (WB) varied by individual, ranging from 117×109/L to 337×109/L (mean, 192±50×109/L). The platelet count was significantly higher in the PRP samples than in native blood (P<0.001 by paired t-test), with an average 6-fold increase; we noted a relatively high degree of variability among individuals (mean, 1,166±402×109/L; range, from 400×109/L to 2,505×109/L; Table 1). The Pearson's correlation between the WB and PRP platelet count was 0.673 (P<0.001). The platelet counts were not correlated with donor age (P=0.222 for WB platelet count; P=0.685 for PRP platelet count).
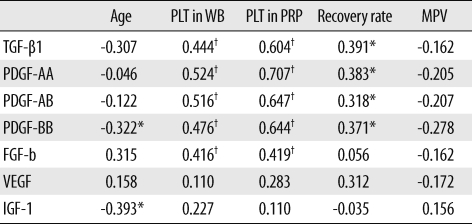
The GF concentrations, with the exception of IGF-1, were significantly higher in aPRPs than in aPPPs (Fig. 2). The concentration of GFs also displayed considerable variation across donors, and the concentrations of TGF-β1, PDGFs, and FGF-b were correlated with the platelet count in both WB and PRP. The concentrations of PDGF-BB and IGF-1 displayed weak negative correlation with age (Table 2).
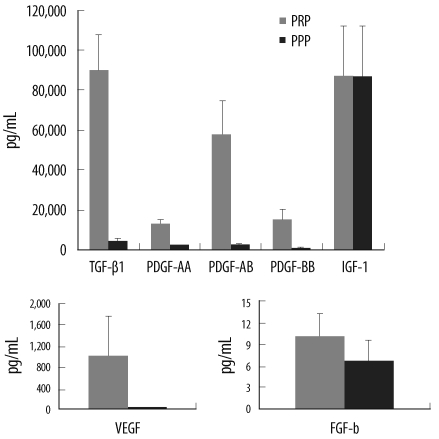 | Fig. 2Quantification of growth factors in activated PRP and activated PPP (P<0.05 between PRP and PPP, except for IGF-1).
Abbreviations: TGF-β1, transforming growth factor-β1; PDGF, platelet-derived growth factor; IGF-1, insulin-like growth factor-1; FGF-b, fibroblast growth factor-basic; VEGF, vascular endothelial growth factor.
|
3. Cluster analysis of 7 GFs and between-group differences in DNA levels or ALP activity in HMSCs
We performed a cluster analysis using the K-means method to identify the variations in GF release patterns and their influences on the biologic effects. Five characteristic release patterns of aPRP GFs were identified and significant differences in PDGF-AA, PDGF-AB, PDGF-BB, TGF-β1 and IGF-1 were noted between the groups. Groups 1 and 3 showed higher concentrations of TGF-β1 and PDGFs, whereas groups 2 and 4 exhibited lower concentrations of TGF-β1 and PDGFs. Groups 1 and 2 showed higher concentrations of IGF-1, whereas groups 3 and 4 showed lower concentrations of IGF-1. Group 5 exhibited substantially lower concentrations of TGF-β1, PDGFs, and FGF-b; however, the group was excluded from ANOVA because its sample size was only 1. The DNA levels differed significantly between groups (P=0.007 by ANOVA) and were higher in groups 1 and 3 than in group 4. The differences in ALP activity between cluster groups were not significant (P=0.289 by K-W test; Table 3).
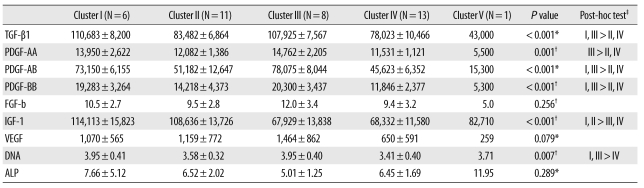
Table 3
Cluster characteristics of growth factors and biologic effects
Values are mean±standard deviation.
Units: growth factors, pg/mL; DNA, µg/well; ALP, nM pNP/min/µgDNA.
P values were determined by *Kruskal-Wallis test, or by †one-way ANOVA, ‡Post-hoc tests were conducted using the Bonferroni correction for the Kruskal-Wallis test or the Scheffe method for ANOVA.
Abbreviations: TGF-β1, transforming growth factor-β1; PDGF, platelet-derived growth factor; FGF-b, fibroblast growth factor-basic; IGF-1, insulin-like growth factor-1; VEGF, vascular endothelial growth factor; ALP, alkaline phosphatase.
![]()
4. Relationship between DNA content or ALP activity and aPRP GF concentrations
The amount of DNA obtained from HMSCs that were cultured for 12 days with 10% aPRPs varied by donor, ranging from 2.59 to 4.59 µg/well (mean, 3.66±0.43 µg/well). The ALP activity per cell also showed considerable individual variations, ranging from 2.83 to 17.96 nM pNP/min/µg DNA (mean, 6.50±2.68 nM pNP/min/µg DNA).
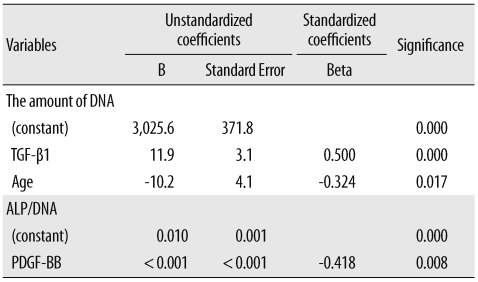
Stepwise multiple regression analysis was employed to identify the independent variables that affected DNA levels and ALP activity. The variables included were donor age, the WB and PRP platelet count, the platelet recovery rate, and aPRP concentrations of the 7 GFs. The amount of DNA was correlated positively with the TGF-β1 concentration (Beta=0.500, P<0.001) and negatively with donor age (Beta=-0.324, P=0.017; Table 4). The ALP activity (ALP/DNA) was correlated negatively with the concentration of PDGF-BB (Beta=-0.418, P=0.008; Table 4).
Go to :
DISCUSSION
Despite 2 decades of clinical study of PRP, little basic data is available regarding the effects of variations in GF levels on biologic function. Because PRP displays the unique characteristics of a mixture of multiple GFs in various concentrations, and because the regenerative potency of PRP probably depends on its GF levels, knowledge of the relationships between GF levels and biologic effects is required to ensure the reliable and reproducible use of PRP [8]. Therefore, in this study, we attempted to explore the relationship between GF levels and biologic effects.
Obtaining a sufficiently large volume of blood from the single donor in order to analyze the various effects was difficult; thus, we employed DNA analysis for proliferation markers and ALP activity for osteogenic markers of HMSCs. First, we cultured HMSCs at several aPRP concentrations (1%, 3%, 10%, and 30%) from 3 donors to evaluate its effects on proliferation and ALP activity, as well as to determine the optimal concentration in our experimental setting. Simultaneously, HMSCs were cultured with the same concentrations of aPPPs in an effort to demonstrate that the effects were attributable to the presence of platelets. Then, HMSCs were cultured with 10% aPRPs from another 39 donors to assess the relationship between the biologic effects and GF concentrations.
Our findings indicated that aPRP induced HMSC proliferation in a dose-dependent manner. The addition of 10% aPRP to the culture medium induced marked cell proliferation in vitro; this result was consistent with the findings of previous studies [13, 14]. Because higher concentration (30%) of aPRP did not promote proliferation, as compared to controls, 10% aPRP may be optimal for the experimental ex vivo expansion of HMSCs. Some studies also showed that a high concentration of PRP did not promote or that it even suppressed cell proliferation; optimal concentration varied with the cell type or preparation method [15-17]. Few studies have suggested the presence of negative regulators in PRP, such as thrombospondin [17], but the reason for the antiproliferative effect of high-concentration-PRP is not clear. Although aPPP could promote cell proliferation when it is activated, the proliferative effect of aPPP was significantly lower than that of aPRP in this and some other studies. Non-activated PPP cannot induce cell proliferation; therefore, the proliferative effect of aPPP also might be attributed to platelet-derived factors [16, 18]. However, why higher concentrations of aPPP suppress HMSCs is still unclear.
In the present study, aPRP suppressed the ALP activity of MSCs. Some studies have reported similar results in that PRP increased migration and proliferation, but reduced the osteogenic differentiation of bone marrow-derived MSCs in vitro [14, 19]. However, in this study, aPPP did not suppress ALP activity at any concentration; therefore, these inhibitory effects may be associated with substances that are derived from platelets.
Substantial variations in the PRP and WB platelet counts were noted. The PRP platelet count was correlated with both donor WB platelet counts and recovery rates. The GF concentrations also showed considerable between-donor variations. The aPRP GF concentrations, except for IGF-1, were significantly higher than the aPPP GF concentrations. This suggested that TGF-β1, PDGFs, FGF-b, and VEGF were derived from platelets, but that IGF-1 did not originate from platelets [20]. The concentrations of TGF-β1, PDGFs, and FGF-b were correlated with the WB and PRP platelet counts, more strongly with the PRP than WB platelet counts. Weibrich et al. [9] noted that the GFs contents were not well correlated with either the WB or PRP platelet counts; however, this result may be attributed to the use of non-activated PRP in their study. In our study, PDGF-BB and IGF-1 concentration were negatively correlated with age, indicating that GFs vary between individuals according to the differences in platelet count and donor age.
Since the efficacy of PRP is based on the production and release of multiple GFs when platelets are activated, we hypothesized that differing GF concentrations might result in different effects on MSC proliferation or osteogenic differentiation. To verify this hypothesis, HMSCs from 39 donors were cultured with 10% aPRP, and the relationships between GF concentrations and DNA contents or ALP activity were evaluated. The individual variations in PRP GF release patterns were noted, and could be divided into 5 groups based on cluster analysis.
The levels of TGF-β1, PDGFs, and IGF-1 differed between the groups. VEGF and FGF-b displayed tendencies similar to those of TGF-β1 and PDGFs, but these differences were not significant. IGF-1 showed a different tendency, suggesting that IGF-1 originated from a different source than the other GFs. The DNA content of HMSCs is significantly higher in groups with higher TGF-β1 and PDGF contents. These results indicated that individual characteristic release patterns of aPRP GFs may be attributable to GFs derived from platelets and IGF-1 and may influence the different proliferative effects.
We then performed multiple regression analysis to evaluate independent factors that could affect DNA content or ALP activity. Multiple regression analysis results showed that the quantity of DNA was positively correlated with the concentration of TGF-β1 and was negatively correlated with donor age. This finding indicates that TGF-β1 is the most mitogenic of the aPRP GFs. TGF-β1 has been shown to stimulate the proliferation of undifferentiated MSCs, and its effects vary with the amount applied [21]. Previous reports have suggested that PDGF-BB and FGF could stimulate the proliferation of MSCs or osteoblastic cells at a concentration of 5 ng/mL [22, 23] and that the concentrations of these GFs in 10% aPRP were substantially lower than 5 ng/mL. The lack of a detectable effect of PDGF-BB and FGF on MSC proliferation in this study might have been the result of low concentrations of PDGF-BB and FGF that might be present in 10% aPRP.
Although the cellular ALP activity did not differ among groups derived from cluster analysis, aPRP inhibited the ALP activity separately from aPPP. Thus, an unknown substance released from the platelets has been hypothesized to suppress osteogenic differentiation. Multiple regression analysis showed that the ALP activity was negatively correlated with the concentration of PDGF-BB in our study. Tokunaga et al. [24] noted that the depletion of PDGFR-β in MSCs attenuated mitogenic and migratory responses, but enhanced osteogenic differentiation. Additionally, Ranly reported that PDGF-BB inhibited demineralized bone matrix-induced intramuscular cartilage and bone formation in a dose-dependent manner [7]. These findings, coupled with the results of our study, indicate that the suppression of ALP activity by aPRP is attributable to PDGF-BB. However, the inhibitory effects of PRP on osteogenesis remain somewhat unclear. The principal disadvantage of this study was that only ALP activity was used as an osteogenic marker, primarily because of the limited available volume of PRP. Kanno et al. [25] demonstrated that ALP activity was suppressed during cell growth, but was enhanced when the cells achieved confluence. Further studies for optimal duration and the effect on other osteogenic markers, such as osteopontin or RUNX2, should be conducted to demonstrate the osteogenic effects of PRP.
In conclusion, aPRPs stimulate MSC proliferation in a dose dependent manner, and they suppress ALP activity during cell proliferation. The GF concentrations reveal individual variations that may result in different biologic effects of aPRP. MSC proliferation was positively correlated with TGF-β1 concentration, and ALP activity was negatively correlated with PDGF-BB concentration in aPRP. These results indicate that individual differences in GFs should be taken into consideration in order to ensure reliable interpretation of the biologic function and standardized application of PRP.
Go to :
 XML Download
XML Download