

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
With the development of bladder outlet obstruction (BOO), the bladder undergoes not only morphologic and physiologic changes, but also functional and symptomatic changes. Despite the high prevalence of BOO among older men, the underlying mechanisms that are responsible for the bladder dysfunction induced by BOO remain poorly understood.
The urothelium was previously considered to be a simple passive barrier between the urinary tract and urine. Recently the urothelium and suburothelial space have received renewed interest because of their possible roles in the pathophysiology of bladder dysfunction. The urothelium is now understood to be an active organ that plays an important role in regulating bladder disorders [1]. It has been shown in some animal models that water transfer can occur across the urothelium from the bladder to the systemic circulation [2-4]. Those studies suggested that the urothelium can mediate water and solute transport under certain circumstances [5-9]. Several studies have shown that rabbit and guinea pig urinary bladder epithelium contains an aldosterone-stimulated sodium transporter [10]. This transporter has been identified as an epithelial sodium channel (ENaC) [11] that is known to be responsible for salt and fluid transport across the epithelia of many tissues [12].
Currently, only limited data are available about AQPs in the mammalian urinary bladder. Spector et al. recently reported on the localization of AQP1, 2, and 3 in rat urothelium [13]. They reported the presence and regulation of AQPs in the urothelium of rats after the rats underwent 24 hours of dehydration and then a water loading test. They suggested that the AQPs in the urothelium may play a role in epithelial cell volume and in controlling osmolarity, and they strongly suggested the possibility of bulk water movement across the urothelium. Rubenwolf et al [14] demonstrated that AQPs are expressed in cultured urothelium and they suggested a potential role for AQPs in transurothelial water and solute transport in humans. However, there are currently no studies that have investigated the expression of AQP in the urothelium of rats after BOO or the functional activity of these proteins in response to BOO. AQP1 is expressed widely in mammalian epithelial and endothelial plasma membranes. Primarily, AQP1 functions as a passive water pore that increases water transport across cell membranes in response to an osmotic driving force. Furthermore, some evidence suggests that AQP1 has an ion channel function that has potential significance to basic and clinical research involving the regulated control of water and ion fluxes across membranes [15]. On the basis of these data, we hypothesized that AQP1 channels may be further impacted by BOO, which may cause bladder dysfunction, at least in part, by an increase in the urothelial permeability of ion and water.
The purpose of this study was to investigate the impact of BOO on AQP1 expression in the rat urinary bladder.
MATERIALS AND METHODS
1. Experimental model
Twenty female Sprague-Dawley rats (230-240 g each) were divided into 2 groups: the control (Con) group (n=10) and the BOO group (n=10). The BOO group underwent partial BOO. The animals were premedicated with xylazine (2.2 mg/kg, IM) and anesthetized with a zolazepam/tiletamine cocktail (4.4 mg/kg, IM). A midline abdominal incision was made and the bladder and proximal urethra were dissected free of the surrounding tissue. To create an intravesical obstruction, a PE-90 (polyethylene catheter-90) catheter was placed beside the proximal urethra and a 3-zero silk ligature was tied around the urethra and catheter. The catheter was subsequently removed and the abdominal incision was closed. The control group underwent a sham operation. Histologic studies were performed on both groups after three weeks. All the rats were in an overnight fasting state before the experiments were performed; they were given a standard diet before that time. The protocol for the animal surgery was approved by the Ethics Committee of the Chonnam National University Medical School.
2. Western blotting
The tissue homogenates (30 µg of protein) were separated by 12% SDS-polyacrylamide gel electrophoresis and the proteins were then transferred to a PVDF membrane (Amersham Pharmacia Biotech, Little Chalfont, United Kingdom). The blots were then washed with Tris-buffered saline Tween-20 (10 mM Tris-HCl, pH 7.6, 150 mM NaCl, 0.05% Tween-20). The membrane was blocked with 5% skimmed milk for 1 hour and was then incubated with the appropriate primary antibody. Monoclonal rabbit antibodies for AQP1 (Chemicon, Temecula, CA, USA) and monoclonal rabbit antibodies for glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (Sigma, St. Louis, MO, USA) were used. The membrane was then washed and the antibody reactions were detected by using goat anti-rabbit-IgG conjugated to horseradish peroxidase. The antibody incubations were performed in a 4℃ incubator. The bands were visualized by using enhanced chemiluminescence (Amersham Pharmacia Biotech). GAPDH was used as an internal control. Densitometry analysis was performed with a studio Star Scanner and by using the NIH image V1-57 software.
3. Immunohistochemistry
The bladder tissue was dissected away from both lateral walls of the bladder. The tissue was placed in 4% paraformaldehyde fixative for 16 hours and the tissue was then processed for washing and dehydration. The tissue was routinely embedded in paraffin, and 6 µm sections were prepared. The tissues were stained with H&E. Immunohistochemistry was performed by using an immunoperoxidase procedure (Vector ABC Kit; Vector Laboratories, Burlingame, CA, USA). The tissue sections were deparaffinized in xylene, dehydrated in a graded series of ethanol solutions, rinsed twice in phosphate-buffered saline (PBS), and then treated with 3% H2O2 in 60% methanol for 30 minutes to quench the endogenous peroxidase activity. After washing twice (5 minutes each time) in PBS, the sections were next incubated for 12 to 14 hours with purified rabbit antibodies for AQP1 (Chemicon) in PBS with 0.3% bovine serum albumin. For the negative control, the sections were incubated in PBS containing only 5% normal goat serum.
4. Immunofluorescence staining
The tissue sections were deparaffinized in xylene, dehydrated in a graded series of ethanol solutions, rinsed in PBS, and then treated with normal goat serum for 30 minutes to block any nonspecific binding. After washing in PBS, the sections were incubated with antibodies for AQP1 (Chemicon) in PBS with 0.3% bovine serum albumin for 12 to 14 hours at 4℃. For a negative control, the sections were incubated in PBS containing only 5% normal goat serum. The sections were then rinsed in PBS and incubated for 30 minutes with the antirabbit IgG conjugated to fluorescein (Vector Laboratories). Finally, the tissue sections were examined and photographed under a fluorescence microscope. The sections were then rinsed 3 times in PBS and incubated sequentially with the biotinylated secondary antibody and the ABC reagent, and each incubation was done for 30 minutes. The sections were then incubated for 5 minutes with the peroxidase substrate solution contained in the kit. At last, the tissue sections were examined and photographed under a light microscope.
RESULTS
All the animals survived for three weeks after the creation of a partial BOO. There was no significant difference in body weight between the 2 groups. The weights (mg) of the bladders were significantly greater in the BOO group (640±15.3) than in the control group (125.3±17.6), and this finding supported the proper creation of BOO (p<0.05).
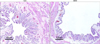
1. Bladder histology and anatomical influence
In the control rats, the urothelium consisted of 3-4 layers of transitional cells and lamina propria. Each microvascular structure was surrounded by scattered smooth muscle bundles and connective tissue. The bladder wall of the BOO animals was thicker than that of the normal controls. In the BOO group, the histology of the bladder tissue showed relative epithelial thinning and distortion of the mucosa compared with that of the control. The proportion of bladder smooth muscle was also markedly higher in the BOO group than in the control group (Fig. 1).
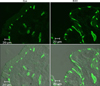
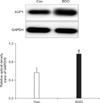
2. The effect of BOO on the expression of AQP1
AQP1 was mainly expressed in the subepithelial capillaries and venules of the lamina propria, including the suburothelial tiny arterioles located just beneath the urothelium. The immunohistochemistry revealed that in terms of the cellular patterns of labeling, the expression of AQP1 in the control group was similar to that in the BOO group. However, the immunohistochemistry (Fig. 2) and immunofluorescence staining (Fig. 3) showed a marked increase in the expression of AQP1 in the BOO group. Western blot analysis revealed bands for AQP proteins at 28 kDa (Fig. 4). AQP1 protein was recognized in all groups, but there was a significant increase in AQP1 protein in the BOO group compared with the control group (p<0.05) (Fig. 4).
DISCUSSION
In the present study, AQP1 was significantly increased in the BOO group. These results suggest that AQPs may play a role in the bladder dysfunction induced by BOO. This study sheds light on the possible occurrence of water and solute movement via AQPs in the rat urinary bladder and the related changes after BOO.
The incidence of overactive bladder symptoms such as frequency, urgency, and urge incontinence in men older than 40 years is estimated to be 16.6% [16]. Symptoms of an overactive bladder can be the result of detrusor overactivity or detrusor instability that is induced by a variety of causes. Detrusor overactivity commonly occurs in association with BOO. Detrusor overactivity is thought to be caused by increased afferent nerve activity, but little is known about the underlying mechanisms by which BOO increases the afferent nerve activity [17]. The urothelium is known to have very low permeability to several urinary solutes and substrates. It has been found that the composition of the urine changes during its transport through the urine passages from the renal pelvis to the urinary bladder, and this finding has been ascribed to a modifying function of the urothelium [18,19]. Hibernating bears that can reabsorb their entire daily urine production during the months of winter are a dramatic example of net urothelial water and solute transport [4]. Araki et al. investigated the role of ENaC in the bladder dysfunction of male patients who were clinically diagnosed with BOO [20]. They reported that in patients with BOO, the ENaC expressed in the human urinary bladder epithelium and the expressed levels of ENaC were significantly higher than in patients who did not have BOO. The authors suggested that the ENaC expression in the bladder epithelium might be implicated in the mechanosensory transduction that occurs in the bladder afferent pathways, and this induces detrusor instability by BOO.
AQPs are a family of transmembrane proteins that transport water across the cell membrane [21]. AQPs are expressed in many fluid-containing tissues and they are involved in many physiological mechanisms, including the transportation of transepithelial fluid, the concentration of urine, and the secretion of gland fluid [22]. AQP1 was the first identified member of the family, and it is expressed at a high rate in water-transporting cells, such as those in the proximal tubule and the thin descending limb in the kidney, and in red blood cells [23]. However, AQP1 is also expressed in vascular smooth muscle cells and vascular endothelial cells where net water movement does not appear to be important [24,25]. To support the hypothesis of water movement in the bladder from the subepithelial capillaries to the lumen, this study investigated the expression of AQPs with focus on the AQP1 expression in the urinary bladder. In the current study, AQP1 was mainly expressed in the capillaries, arterioles, and venules of the urinary bladder.
Our findings agree with the results of a previous study by Spector et al, who showed that AQPs in the genitourinary tract urothelium likely play a role in regulating urothelial cell volume and osmolality [13]. They revealed that dehydrated rats had a significant upregulation of AQP expression, and this finding provides presumptive evidence that AQPs are involved in water and solute transport across the urothelium [13]. Rubenwolf et al demonstrated the expression of AQPs in cultured human urothelial cells, and they suggested that AQPs play a potential role in transurothelial water and solute transport in the human bladder [14]. It has been reported that AQP1 is abundant in a variety of tissues, and the tissues that express AQP1 exhibit high water permeability and a high osmotic urine concentration, gall secretion, formation of cerebrospinal fluid, and spermatozoa concentration [26]. Lu et al recently reported that AQP1 is expressed in male reproductive organs such as the rete testis, vas deference, prostate, and seminal vesicle [26], and this finding provides valuable information on the role of AQP1 in water transport to regulate water homeostasis in male reproductive physiology. The rete testis is the initial segment of the male reproductive tract and has a large vascular surface area. The rete testis alters the nature of the fluid produced by the seminiferous tubule for making more homogeneous sperm [27]. AQP1 has been observed in the rete testis and a significant change in water content was found in AQP1 knockout mice, which indicates that this water channel does indeed play an exclusive role in water transport for forming the rete testis fluid and sperm transit [28].
No studies to date have investigated the expression of the AQP family members in the urinary bladder of the BOO rat model or the changes in the functional activity of these proteins in response to BOO. In the present study, the expression of AQP1 was significantly affected by BOO. These findings suggest that AQPs are influenced by the bladder dysfunction induced by anatomical changes such as BOO. One of the possible reasons behind this influence on the expression of AQP is believed to be due to the significance of the location of the AQP expression at the suburothelial space immediately below the basal lamina of the epithelium, which is well supplied by microvasculature (AQP1). Our results may suggest that BOO leads to a significant up-regulation of AQP1 expression, which provides presumptive evidence that AQPs are involved in the bladder dysfunction induced by BOO. This bladder dysfunction probably occurs via modification of the urothelial water and solute composition. The limitation of this study is that we did not fully unveil the precise functional activity of AQP1, although we did show the changes in the expression of AQP1 in the BOO rat urinary bladder. Further studies are needed to investigate the expression and localization of all the AQP family members in the urinary bladder and their functional role in the underlying mechanisms of bladder pathophysiology.
CONCLUSIONS
BOO causes a significantly increased expression of AQP1 in the rat urinary bladder. This may imply that AQPs have a functional role in the bladder dysfunction that occurs in association with BOO. Further study is needed to clarify the exact functional role of the different AQPs in the urinary bladder.

 XML Download
XML Download