

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Osteoclasts
Osteoclasts are unique bone-resorbing cells, and mice carrying a mutant gene responsible for osteoclast formation exhibit osteopetrotic phenotype[1,2]. Osteoclasts are giant cells containing 10~20 nuclei or sometimes more than 100 nuclei in each cell, and the multinucleation is a consequence of fusion of mononuclear osteoclasts. Osteoclasts are derived from hematopoietic stem cells or monocyte/macrophage lineage cells, and an interaction between RANKL (receptor activator of NF-κB ligand) and RANK is required for osteoclast formation[3,4]. RANKL is expressed in osteoblasts whereas RANK is expressed in osteoclast precursor cells. Although RANKL is a transmembrane ligand, a genetically engineered soluble form of RANKL is also effective for osteoclast differentiation. We were able to generate osteoclasts without osteoblastic cells in the presence of M-CSF (macrophage colony stimulating factor) and RANKL[5,6]. This pure culture ystem enabled us to analyze gene expression profiles in osteoclasts.
Identification of osteoclast precursor cells and their differentiation
To establish a pure osteoclast culture system, we tried to isolate osteoclast precursor cells. Bone marrow mononuclear cells were subdivided to three populations based on expressions of c-Fms and RANK; c-Fms+RANK-, c-Fms+RANK+, c-Fms-RANK+. c-Fms is a receptor of M-CSF, whereas RANK is a receptor of RANKL. Each cell population was sorted by FACS and cultured in the presence of M-CSF and RANKL. Osteoclastogenesis was evaluated by tartrate resistant acid phosphatase (TRAP) activity, an enzymatic marker of osteoclasts. Among the populations cultured, c-Fms+RANK-cells had the highest potential to differentiate into osteoclasts, and almost all of the cells expressed TRAP activity[6]. Thus we could establish a pure osteoclast culture system, and decided to use this population in the following experiments. We found that RANK expression was induced in c-Fms+RANK- cells, and the following RANKL stimulation induced effective osteoclast differentiation[5]. Since osteoclastogenesis is strictly limited to bones, some unique factor other than cytokines essential for osteoclast differentiation might be present in the bone. We focused on the hard substrate of bones as an adherent condition. The osteoclastogenesis was significantly inhibited in non-adherent conditions prepared by methylcellulose culture compared with adherent cultures prepared by liquid culture, and the inhibited differentiation in non-adherent conditions was fully restored by transferring those cells into an adherent condition, indicating that osteoclast differentiation is anchorage dependent[6]. Thus we considered that an adherent condition prepared by bones is involved in osteoclastogenesis.
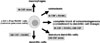
Although osteoclastogenesis was strongly inhibited in non-adherent conditions, the inhibition was not complete. Thus, some other factor might exist inhibiting osteoclast differentiation in the soft tissues. We focused on GM-CSF, and found that osteoclastogenesis induced by M-CSF plus RANKL was completely inhibited by GM-CSF[7]. The osteoclast precursor cells differentiated into TRAP-negative mononuclear cells in the presence of M-CSF, RANKL and GM-CSF. These cells express CD11c, a marker of dendritic cells, indicating that GM-CSF induces the cells to a dendritic cell lineage. The commitment of common precursor cells to macrophages, osteoclasts, and dendritic cells in the presence of specific cytokines are shown in Fig. 1. GM-CSF strongly reduced the expression of c-Fos, an essential transcription factor for osteoclast differentiation, and forced expression of c-Fos rescued the inhibition of osteoclastogenesis by GM-CSF, suggesting that downregulation of c-Fos is critical for the inhibition of osteoclast differentiation by GM-CSF.
Identification of an osteoclast fusion molecule; DC-STAMP
Since cell-cell fusion is mediated through cell contact, various cell surface molecules such as E-cadherin, Macrophage Fusion Receptor (MFR), CD44, CD9 and CD81 have been implicated in the multinucleation of osteoclasts and macrophages[8~11]. However, defects in multinucleation of osteoclasts and macrophages have not been shown by gene targeting. In order to isolate osteoclast cell-cell fusion molecules, we undertook DNA subtraction screens between multinuclear osteoclasts and mononuclear macrophages. As above, multinuclear osteoclasts were induced by M-CSF and RANKL, whereas mononuclear macrophages were induced by M-CSF alone. We have already found that various molecules such as RANK, c-Fms, integrinαvβ3 and Mac-1 are equally expressed in both osteoclasts and macrophages. Thus DNA subtraction screen was considered effective in identifying osteoclast specific molecules. More than 100 molecules were identified by the screen, and DC-STAMP was selected as an osteoclast specific molecule[12].
Characterization of DC-STAMP in osteoclasts
DC-STAMP was identified in dendritic cells as DC -STAMP or IL-4 stimulated macrophages as FIND[13,14]. DC-STAMP was also detected in osteoclasts[15]. In order to understand the role of DC-STAMP in vivo, we generated DC-STAMP deficient mice[12]. The DC-STAMP deficient mice show no growth abnormalities, however, osteoclast cell-cell fusion was completely abrogated in DC-STAMP deficient mice in vivo and in vitro. Osteoclasts in DC-STAMP deficient mice show TRAP-activity, and the osteoclast differentiation markers such as cathepsin K was expressed in DC-STAMP deficient osteoclasts as in wild type osteoclasts. NFATc1 and c-Fos, both of which were essential transcription factors required for osteoclast differentiation, were induced in DC-STAMP deficient osteoclasts at equivalent levels to wild type osteoclasts. Furthermore, the formation of ruffled boarder, a highly specific cytoskeletal feature seen in wild type osteoclasts, was also detected in DC-STAMP-/- osteoclasts. The osteoclast cell-cell fusion was effectively rescued by retro viral transduction of DC-STAMP into DC-STAMP deficient osteoclast progenitor cells, suggesting that DC-STAMP is specifically required for osteoclast cell-cell fusion. In fact, the mononuclear osteoclasts seen in DC-STAMP deficient mice showed bone resorbing activity, and bone marrow cavity formation and teeth eruption were observed in DC-STAMP deficient mice indicating that the osteoclasts in DC-STAMP deficient mice are mononuclear but bone-resorbing cells. However, the bone resorbing activity evaluated by pit formation assay in DC-STAMP deficient osteoclasts was significantly low compared with that of wild type osteoclasts, even though the total number of nuclei was equal. In in vivo analysis, an elevated radio-opacity was seen in DC-STAMP-/- mice by Soft X-ray analysis, and the bone mineral density (BMD) and bone volume per tissue volume (BV/TV) was increased in DC-STAMP-/- mice compared with wild type mice, suggesting that multinucleation enhances the bone resorbing efficiency of osteoclasts. Thus loss of osteoclast cell fusion leads to an increased bone mass.
Characterization of DC-STAMP in FBGCs
Foreign body giant cells (FBGCs) are derived from monocyte/macrophage lineage cells like osteoclasts, and are generated in response to foreign bodies at the site of implantation[16]. Both osteoclasts and FBGCs form multinuclear cells by fusion of mononuclear cells. Interestingly, multinuclear FBGCs formation induced by implantation of foreign bodies was also completely abrogated in DC-STAMP-/- mice. FBGCs are forme in vitro from macrophage and osteoclast common precursor cells by stimulation with several cytokine combinations such as IL-3 plus IL-4[17] or GM-CSF plus IL-4. FBGC formation in vitro was also completely inhibited in DC-STAMP-/- cells, indicating that DC-STAMP is universally required for osteoclast and macrophage cell fusion.
Mechanisms of osteoclast cell fusion via DC-STAMP
DC-STAMP knock-out mice were generated as EGFP knock-in mice, therefore, only mononuclear EGFP-positive cells were induced from DC-STAMP deficient osteoclast precursor cells in the presence of M-CSF plus RANKL. However, multinuclear EGFP-positive cells were induced when osteoclast precursors isolated from DC-STAMP-/- and wild type mice were mixed and cultured together in the presence of M-CSF plus RANKL. This interesting phenomenon suggests that fusion between EGFP-positive DC-STAMP-/- cells and wild type osteoclast precursor cells was induced. These data indicate that a putative DC -STAMP ligand might be expressed on osteoclasts, and the interaction between DC-STAMP and its putative ligand may induce the cell-cell fusion, and therefore the expression of DC-STAMP is not required in all the cells participating in multinucleation. Heterogeneous EGFP expression in DC-STAMP-EGFP knock-in cells during the course of osteoclast differentiation supports this possibility. Thus, DC-STAMP might be involved in cell-cell interactions in a receptor-ligand fashion. Further analysis is required to clarify the mechanisms of cell-cell fusion via DC-STAMP.
 XML Download
XML Download