

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Activation of inflammation by metabolic stress in diabetes is associated with insulin deficiency caused by destruction of pancreatic beta cells and insulin resistance in adipose tissue and liver [1,2]. Adipocytes, which secrete several adipokines (leptin, resistin, tumor-necrosis factor [TNF]-α, interleukin [IL]-6, and PAI-1), induce insulin resistance and inflammation throughout the entire body through a decrease in adiponectin [3]. The secretion of cytokines caused by the activation of NF-κB also induces insulin resistance in fatty liver [4].
Some studies have reported that TNF-α is positively associated with insulin resistance, which is accompanied by diabetes and obesity [5,6]. In some studies that investigated the relationship between insulin resistance and TNF-α, increases in TNF-α were found in the skeletal muscle of patients who had a high insulin resistance [7]. In addition, TNF-α inhibits activation of the insulin receptor tyrosine kinase by blocking insulin receptor substrate (IRS) phosphorylation and Akt substrate 160 phosphorylation, which are important proteins in the insulin signaling pathway in skeletal muscle [8-10]. Based on these results, it seems that an increase in TNF-α is not just a byproduct of inflammation, but may directly influence inflammation of skeletal muscle.
Thiazolidinedione (TZD) is an insulin sensitizer that functions not only to enhance the effect of insulin by controlling gene transcription through binding with nuclear receptor peroxisome proliferator activated receptor (PPAR)-γ, but also to inhibit inflammation of adipose tissue, immunologic cells and blood vessels through several inflammatory pathways (STAT, AP-1 and NF-κB pathway) [11-13].
In this study, we investigated the role of skeletal muscle inflammation in the pathogenesis of insulin resistance in type 2 diabetes in the Otsuka Long Evans Tokushima Fatty (OLETF) rat, a well-known type 2 diabetes animal model. OLETF rats progress to hyperglycemia in postnatal week 18 and fibrosis of pancreatic beta cells in postnatal week 40 [14]. We also investigated whether the anti-inflammatory effect of rosiglitazone, which has been shown in adipose tissue and inflammatory cells, is also present in skeletal muscle.
METHODS
Laboratory animals
Five-week-old OLETF rats (n = 21) and Long-Evans Tokushima Otsuka (LETO) rats (n = 11) were obtained from the Otsuka Pharmaceutical Co. Ltd. (Tokushima, Japan). Each rat was housed in a separate cage and was fed animal feed. The cages were maintained at a constant temperature of 22 ± 2℃ and the light-dark cycle was adjusted automatically every 12 hours. To extract tissues before the experiment, 5 rats from the OLETF group and 5 rats from the LETO group were sacrificed. Between the 28th and 40th weeks, LETO rats (n = 6) were fed normally, and OLETF rats were divided into a non-treated group (n = 8) and a rosiglitazone-treated (GlaxoSmithKline Pharmaceuticals, Philadelphia, PA, USA) group (3 mg/kg body weight, n = 8) (Fig. 1). At the 40th week, weight and urine glucose levels were measured, and the rats were sacrificed to collect gastrocnemius muscle samples.
The body weights of the animals were measured every 4 days at 4 o'clock in the afternoon between the 8th and 40th weeks.
Serologic tests
At the 28th and 40th weeks, fasting blood was extracted from the hearts of OLETF and LETO rats. Plasma was separated from whole blood and refrigerated at -70℃. Serum glucose level was measured by glucose oxidase reaction method using an enzyme reagent (Asan Pharmaceutical Co., Seoul, Korea). The concentration of insulin was measured by enzyme-linked immunosorbent assay (ELISA) kit (Shibayagi Co., Ishihara, Japan). The concentration of free fatty acids was measured by the kinetic alkaline picrate method using an enzyme reaction kit (Asan Pharmaceutical Co.). Homeostasis model assessment (HOMA)-insulin resistance (IR) and HOMA-β (HOMA beta cell function) were estimated using the following formula: [Fasting insulin (µIU/mL) × fasting plasma glucose (mmol/L)]/22.5; HOMA-β = 20℃ fasting insulin (µU/mL)/fasting glucose (mg/dL) - 63 [15].
Measurement of inflammatory cytokines
At the 28th and 40th weeks, the concentrations of TNF-α, IL-1β and IL-6 were measured from the sampled blood plasma using an ELISA kit (eBioscience, San Diego, CA, USA).
Expression of inflammatory cytokine genes
To extract gastrocnemius muscle samples, the rats were sacrificed by intraperitioneal injection of 2.5% tribromoethanol at the 28th and 40th weeks. The extracted gastrocnemius samples were immediately frozen with liquid nitrogen and ground with a homogenizer. Total RNA was separated from the ground tissues using TRIzol reagent. The cDNA was synthesized using oligo-(dT) primer and AccuScript™ high fidelity 1ST strand cDNA synthesis kit (Stratagene) at 65℃ for 5 minutes, at 42℃ for 1 hour and at 70℃ for 15 minutes. For the PCR reaction, 10 × PCR buffer (10 mM Tris-HCl, pH 8.3, 50 mM KCl, 0.1% Triton X-100), 250 µM dNTP mix, 1 U Taq polymerase (Takara) and a specific primer for each cytokine were mixed with 1 µg cDNA. The reaction was incubated at 94℃ for 30 seconds for denaturation, at 57-62℃ for 30 seconds for annealing, and at 72℃ for 30 seconds for extension. This cycle was repeated 25-35 times. ACTIN was used for the comparison with PCR reaction. The primer sequences for the PCR reaction are shown below.
Actin; Forward, 5'-TCA TGA AGT GTG ACG TTG ACA TCC GT-3'; Reverse, 5'-CCT AGA AGC ATT TGC GGT GCA CGA TG-3', TNF-α ; Forward, 5'-AAA GCA TGA TCC GAG ATG TG-3'; Reverse, 5'-AGC AGG AAT GAG AAG AGG CT-3', IL-6: Forward, 5'-CCG GAG AGG AGA CTT CAC AG-3'; Reverse, 5'-AGA ATT GCC ATT GCA CAA C-3', IL-1β; Forward, 5'-CAT CTT TGA AGA AGA GCC CG-3'; Reverse, 5'-GGG ATT TCG TTG TTG CTT GT-3'.
Inflammatory signal molecules
Ground gastrocnemius muscle liquid, which was extracted from the 28th and 40th week, was washed twice with HBSS buffer, and then lysed on ice with RIPA buffer (50 mM HEPES, pH 7.4, 150 mM NaCl, 1% deoxycholate, 1 mM EDTA, 1 mM PMSF, 1 µg/mL aprotinin). To observe NF-κB expression, the nucleus was extracted using NE-PER nuclear and cytoplasmic extraction reagent (Pierce, Rockford, IL, USA). The protein was measured by the Bradford method. The same quantity (20 µg) of protein was mixed with 2X sample buffer (100 mM Tris-HCl, pH 6.8, 200 mM dithithreitol, 4% SDS, 0.2% bromophenol blue, 20% glycerol). Then, the mixture was separated with 10% SDS-PAGE. After electrophoresis, the protein was transferred to nitrocellulose membrane at 4℃, 30 V for 16 hours. The membrane was blocked with 10% skim milk at room temperature for 1 hour, and then the membrane was reacted for 2 hours at room temperature with a dilution of monoclonal antibody for β-actin, ERK1/2 MAPK or NF-κB (Cell Signaling Technology; #9101, #3036); antibodies were diluted with Tris-buffered saline containing 0.05% tween (TBS-T). The membrane was then washed three times with TBS-T. Then the membrane was reacted with HRP-conjugated anti-rabbit IgG secondary antibody at room temperature for an hour, and bands were visualized with an enhanced chemiluminescence kit and X-ray film (Amersham Pharmacia Biotech, San Francisco, CA, USA).
Statistical analysis
All results are shown as the mean and standard deviation based on three experiments. Statistical analysis of the difference between groups was analyzed by one-way ANOVA (GraphPad Prism program; GraphPad Software Inc., San Diego, CA, USA) and Student t-test. A P value less than 0.05 was considered statistically significant.
RESULTS
Changes in body weight
The weights of the OLETF and LETO rats were measured every 4 weeks between the 18th and 40th week. The weights of both OLETF and LETO rats decreased with time until the 36th week. An insignificant decrease in weight in normally fed OLETF rats was observed at the 40th week. The increase in the weight of the OLETF rats was greater than that of the LETO rats, regardless of rosiglitazone administration. At the 36th week, the average weight of the LETO rats was 534.9 ± 7.7 g, while that of rosiglitazone-treated OLETF rats was 733.9 ± 18.6 g, and that of untreated OLETF rats was 681.1 ± 18.0 g. At the 40th week, the average weight of the LETO rats was 558.3 ± 8.7 g, while that of untreated OLETF rats was 655.7 ± 48.9 g and that of the rosiglitazone-treated OLETF rats increased up to 757.8 ± 19.0 g (Fig. 2). The weight gain of the rosiglitazone-treated OLETF rats was greater than that of the untreated OLETF rats at the 32nd week. However, the difference was not statistically significant until the 40th week.
Changes in serologic markers
To investigate the chemical changes related to the administration of rosiglitazone, blood was obtained from both OLETF and LETO rats at the 28th and 40th week. Concentrations of free fatty acid (FFA), insulin and glucose were measured from the plasma (Fig. 3). At the 28th week, the fasting glucose concentrations of the LETO and OLETF rats were 167.9 ± 40.1 mg/dL and 229.8 ± 44.2 mg/dL, respectively. The glucose level was higher in the OLETF rats than in the LETO rats. At the 40th week, the glucose level of the LETO rats was 164.5 ± 26.8 mg/dL, while that of the rosiglitazone-treated OLETF rats was 224.1 ± 42.5 mg/dL, and that of the untreated OLETF rats was 276.7 ± 44.4 mg/dL. The glucose level of the rosiglitazone-treated OLETF rats decreased significantly compared with that of untreated OLETF rats.
At the 28th week, the insulin concentrations of the LETO rats and OLETF rats were 0.94 ± 0.1 ng/mL and 2.66 ± 0.4 ng/mL, respectively. The insulin concentration of the OLETF rats increased more than that of the LETO rats (P < 0.05). At the 40th week, the insulin concentration of the LETO rats was 1.89 ± 1.6 ng/mL, that of the rosiglitazone-treated OLETF rats was 2.01 ± 1.4 ng/mL and that of the untreated OLETF rats was 1.89 ± 1.6 ng/mL. The insulin concentration decreased significantly in the rosiglitazone-treated group (P < 0.05, Fig. 3).
There were no statistically significant changes in HOMA-β and HOMA-IR among the LETO rats, rosiglitazone-treated and untreated OLETF rats (data not shown). The mean values of HOMA-IR among the LETO rats, rosiglitazone-treated and untreated OLETF rats were 0.8 ± 0.35, 2.4 ± 1.05 and 1.83 ± 0.86, respectively. Insulin resistance increased more in untreated OLETF rats than in LETO rats. There was a trend of decreased insulin resistance in rosiglitazone-treated OLETF rats compared with untreated OLETF rats (data not shown).
At the 28th week, the FFA concentrations of the LETO and OLETF rat groups were 484.8 ± 77.9 mg/dL and 311.8 ± 76.7 mg/dL, respectively (Fig. 3). At the 40th week, the FFA concentration of the LETO rats was 505.3 ± 82.7 mg/dL, that of the untreated OLETF rats was 276.1 ± 18.3 mg/dL and that of the rosiglitazone-treated OLETF rats was 280.1 ± 46.6 mg/dL. At the 40th week, the FFA level was lower in the OLETF group compared to the LETO group. In contrast, a decrease in the FFA level was not observed in the rosiglitazone-treated group compared with the untreated group.
Changes in inflammatory cytokines
Serum TNF-α concentrations at the 40th week were 261.9 ± 8.3 pg/mL in the LETO rats, 613.5 ± 64.3 pg/mL in the untreated OLETF rats, and 282.1 ± 11.84 pg/mL in the rosiglitazone-treated OLETF rats (Fig. 4). The TNF-α concentration of the OLETF rat group was significantly increased compared with that of the LETO rat group at the 40th week. In the OLETF rat group, TNF-α secretion was significantly decreased by rosiglitazone administration.
For IL-1β at the 40th week, the mean concentration in the LETO rat group was significantly lower than that in the untreated OLETF rat group (125.6 ± 7.9 pg/mL vs. 189.8 ± 30.6 pg/mL, respectively). The IL-1β concentration of the rosiglitazone-treated OLETF rat group at the 40th week, was 137.8 ± 5.7 pg/mL, which was significantly lower than that of the untreated group (Fig. 4).
For IL-6 at the 40th week, the mean concentration in the LETO rat group was significantly lower than that of the untreated OLETF rat group (261.8 ± 19.5 pg/mL vs. 906.5 ± 49.1 pg/mL, respectively). The rosiglitazone-treated rats had significantly lower IL-6 levels (709.1 ± 61.2 pg/mL) compared with untreated rats (Fig. 4).
Changes of inflammatory cytokine gene expression in skeletal muscle
The gene expression of inflammatory cytokines in the OLETF rat group was higher than that of the LETO rat group (Fig. 5). Cytokine gene expression was lower in the rosiglitazone-treated OLETF rat group than in the untreated OLETF rat group, although the difference was not statistically significant.
Changes in inflammatory signal molecules expression
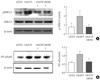
To investigate the effects of rosiglitazone administration on the inflammatory pathway, expression of signaling molecules in ERK1/2 MAPK and NF-κB pathways was examined in skeletal muscle tissue of LETO and OLETF rats by Western blot (Fig. 6). The phosphorylation of ERK1/2 MAPK of the LETO rats was shown to increase compared with that in the OLETF rats. The phosphorylation of ERK1/2 was lowered by rosiglitazone treatment in OLETF rats (Fig. 6A). In addition, for NF-κB, there was an increase in the expression of the NF-κB p65 subunit in the nucleus in OLETF rats, and this was decreased by administration of rosiglitazone (Fig. 6B).
DISCUSSION
Skeletal muscle mainly uses glucose as an energy fuel. Insulin resistance of muscle, caused by accumulation of fatty acid and reduced expression of glucose transporter, plays a significant role in the pathogenesis of type 2 diabetes. Normal muscle tissues are not involved in the inflammatory process, except for in the condition of muscle atrophy, which is seen in cachexia or chronic inflammation of muscles. Human leukocyte antigen (HLA) class I, HLA class II and secretion of inflammatory cytokine increase when a proinflammatory stimulus is given to normal muscle tissues [15]. TNF-α mRNA expression after delivery was higher in gestational diabetes patients than in patients with normal glucose tolerance, and this was also related to insulin resistance [16]. The inflammatory process was observed in the skeletal muscle of diabetes patients, in whom chronic systemic inflammation is known to be present. However, the degree of inflammation of skeletal muscle in relation to insulin resistance in diabetes has never been shown. An increase in adipose tissue caused by obesity activates the JNK and NF-κB pathway, and also induces inflammation in liver through secretion of several inflammatory cytokines. This phenomenon is known to contribute to hyperglycemia through inappropriate inhibition of gluconeogenesis in liver [4]. Our hypothesis was that increased inflammation in skeletal muscle tissues of diabetes patients contributes to increases in insulin resistance in type 2 diabetes, just as there is inflammation in liver tissues in diabetes patients. Then, we examined inflammation markers in skeletal muscle tissue and plasma in OLETF rats, which is an animal model of type 2 diabetes.
In this experiment, at the 40th week, gene expression of inflammatory cytokines (TNF-α, IL-1β and IL-6) and expression of ERK1/2 MAPK and NF-κB in the gastrocnemius muscle was increased in OLETF rats compared to LETO rats. Increased expression of the transcriptional factors of inflammatory pathways such as ERK1/2 MAPK and IκB/NF-κB in the nucleus is known to be involved in the pathogenesis of insulin resistance in skeletal muscle. There have been other studies that observed activation of NF-κB caused by reduction of inhibitor of κBβ (IκB) at skeletal muscle of type 2 diabetes patients is related with insulin resistance [17]. Activation of NF-κB in adipose tissue, which is caused by TNF-α stimulus, induces insulin resistance through inhibition of insulin signaling [18,19].
The mode of action of TZD has not verified completely, but it is known to bind with PPAR-γ selectively in adipose tissue, muscle tissue and liver tissue, which are the target organs of insulin action. It accelerates gene expression of glucose transporter (GLUT)-1, GLUT-4 and activates differentiation of adipose tissue and inhibits expression of TNF-α, hepatic glucokinase. Finally, it improves insulin resistance and recovers glucose homeostasis in the human body. In this experiment, we found that rosiglitazone plays a role in reducing glucose and insulin levels among various biochemical markers. We observed an improvement in insulin resistance, although there was not an increase in insulin secretion ability. Rosiglitazone-treated OLETF rats gained their weight gradually until the 40th week, while untreated OLETF rats lost weight after the 40th week, which is considered to be a negative side effect of rosiglitazone.
It is known that TZDs have an anti-inflammatory effect through inhibition of the transcription process in the inflammation pathway. PPAR-γ expression was abundantly found in adipose tissue, macrophages and endothelial cells of blood vessels and it was also reported that TZDs decrease NF-κB expression at colonic epithelial cells in inflammatory bowel disease [20]. However, its anti-inflammatory effect on skeletal muscle, which is the target organ of insulin action, has not been clarified yet. It is known that rosiglitazone has an effect on several inflammatory pathways, such as STAT, AP-1, and NF-κB [21]. Rosiglitazone inhibits inflammatory cytokine secretion at monocytes and macrophages by interruption of NF-κB in these inflammatory pathways [11-13]. In our study, at the 40th week, inflammatory cytokines (TNF-α, IL-1β and IL-6) were reduced significantly in rosiglitazone-treated OLETF rats in comparison to untreated OLETF rats. The expression of inflammatory cytokine mRNA (TNF-α, IL-1β and IL-6) was found to be decreased in rosiglitazone-treated OLETF rats. The inflammatory pathways of ERK/MAPK and NF-κB were inhibited at skeletal muscle of rosiglitazone-treated OLETF rats. This study result shows that rosiglitazone, which increases the expression of glucose transporters and to decrease the accumulation of lipids at skeletal muscle [22,23], also interrupts the expression of inflammatory cytokine (TNF-α, IL-1β and IL-6) mRNA and thus contributes to the inhibition of inflammatory pathway of ERK/MAPK and NF-κB at skeletal muscle [24,25]. However, there is still doubt that only inhibition of NF-κB pathway at skeletal muscle is sufficient to improve insulin resistance in the whole body, because a previous study showed an opposite result based on an experiment with muscle-specific IKK2 (IκB kinase) knockout mice [26]. The anti-inflammatory effect of rosiglitazone on insulin resistance must be studied more extensively through the measurement of GLUT-4 expression, glucose receptor and IRS-1 in the skeletal muscle of OLETF rats. In addition, it is necessary to investigate whether Rel translocation, Rel DNA binding and IκBα degradation, which are considered to be involved in the pathogenetic mechanisms that inhibits NF-κB pathway, are related to the anti-inflammatory effect of rosiglitazone.
In this experiment, we did not observe improvement in FFA level, which is a well-known effect of rosiglitazone in other tissues. Although the weight of OLETF rats increased, the FFA level did not increase in the OLETF rats compared to the LETO rats between the 28th and 40th week. Thus, we were unable to detect an improvement in lipids. However, the anti-inflammatory effect of rosiglitazone was observed. It has already been shown that low dose rosiglitazone exerts an anti-inflammatory effect independently of improvements in lipid and insulin resistance in type 2 diabetes [27].
In this study, we observed the relationship between changes in skeletal muscle inflammation and chronic inflammation in hyperglycemic status. Rosiglitazone directly inhibited the inflammatory process in the skeletal muscle in our rat model, which is considered to be a component of the pathogenesis of insulin resistance in skeletal muscle. These effects might help to improve insulin resistance, not only in skeletal muscle, but also in the entire body.





 XML Download
XML Download