

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Idiopathic pulmonary fibrosis (IPF) is a progressive, lethal lung disease characterized by alveolar epithelial cell injury, proliferation of activated fibroblasts and myofibroblasts and extracellular matrix accumulation/remodeling that leads to irreversible distortion of the lung parenchyma12.
It has been suggested that the origin of the activated fibroblasts and myofibroblasts are from several, but not mutually exclusive, pathways. Traditionally, migration/proliferation of the resident fibroblasts is thought to be a major source of activated fibroblasts and myofibroblasts3. Recently, it has been shown that activated fibroblasts and myofibroblasts could arise from other cells like bone marrow-derived circulating fibrocytes4, microvascular pericytes5, endothelial cells6, and alveolar epithelial cells (AECs)78.
Epithelial-to-mesenchymal transition (EMT) is highlighted as an important, possible mechanism of fibrosis within various organs7910. There is evidence in which cuboidal epithelial cells, originally possessing the epithelial cell marker, are transformed to fibroblast-like spindle cell morphology with new expression of the mesenchymal cell marker after transforming growth factor β1 (TGF-β1) treatment, and cells co-expressing both the epithelial and mesenchymal markers are seen in the human IPF lung as well as TGF-β1–treated AECs911. The histological evidence implies that EMT is a functional transition of polarized epithelial cells into migratory mesenchymal cells, suggesting that the origin of the activated fibroblasts and myofibroblasts are the epithelial cells that are injured and follow an aberrant healing process, losing their normal epithelial regenerative capacity9.
Injured epithelial cells prompt the fibrogenic process by releasing TGF-β1, which is a prototypical profibrotic growth factor and well known for having a pivotal role in inducing EMT, commonly throughout all organs10. TGF-β1 expression is up-regulated in the IPF patient's lungs, especially in bronchiolar and AECs, and the extent of adjacent subepithelial TGF-β1 deposition is associated with fibroblast proliferation/accumulation 12.
Currently, there are no proven drugs that cure IPF. Antiinflammatory drugs such as azathioprine and prednisone have no effect on slowing the progression of lung fibrosis and show no survival benefit1314. Meanwhile, anti-fibrotic agents such as nintedanib and pirfenidone have been proven to slow progression of IPF14.
Recently, we reported that apolipoprotein A1 (ApoA1) has an anti-inflammatory and anti-fibrotic effect on the bleomycin and silica-induced lung fibrosis model1516. Lung ApoA1 expression is decreased in IPF as well as in the bleomycininduced fibrotic mouse, and intranasal ApoA1 treatment has been shown to attenuate lung inflammation and fibrosis in the mouse model15. In sequence, we also demonstrated that ApoA1 overexpression attenuated silica-induced established lung fibrosis by using an ApoA1 transgenic mouse model16. Although, our previous data represented anti-inflammatory and anti-fibrotic activities of ApoA1, we could not evaluate the precise mechanism of its anti-fibrotic activities, especially in EMT.
In this study, we have investigated whether ApoA1 modulates TGF-β1–induced EMT and have explored possible mechanisms using the AEC line and the silica-induced lung fibrosis mouse model.
Materials and Methods
1. TGF-β1–induced EMT using A549 cells
A549 cells (ATCC CCL185; American Type Culture Collection, Manassas, VA, USA) were obtained from the ATCC and maintained in Ham's F12K medium with 10% fetal bovine serum, 100 U/mL penicillin, and 100 mg/mL streptomycin. To induce EMT, cells were stimulated for 48 hours in serum free medium (SFM) with 0.1% bovine serum albumin (BSA) and TGF-β1 (5 ng/mL, R&D Systems, Minneapolis, MN, USA). To evaluate the effect of ApoA1, the culture medium was replaced with SFM supplemented with 0.1% BSA, TGF-β1 (5 ng/mL) and recombinant human ApoA1 (100 ng/mL, Calbiochem, San Diego, CA, USA) and the cells were incubated for 48 hours. Control cells were maintained in SFM with 0.1% BSA for 48 hours. The cellular morphological change was evaluated using phase control microscopy (Carl Zeiss Microsystems, Thornwood, NY, USA). Western blot and immunofluorescence were used to measure the expression of N-cadherin, E-cadherin, and α-smooth muscle actin (α-SMA). Smad and non-Smad pathway related molecules and TGF-β1 receptor protein (type I [TβRI] and II [TβRII]) expression were measured by western blot in the cell.
2. Silica-induced lung fibrosis mouse model
Male ApoA1 overexpressing transgenic mice (6–8 weeks old) were treated with silica intratracheally following our previously described method16. On day 0, the transgenic mice received 20 mg of sterile silica crystals (median diameter, 1–5 µm; Sigma-Aldrich, St. Louis, MO, USA) in endotoxin-free water in a total volume of 100 µL by intratracheal delivery. ApoA1 overexpressing transgenic and ApoA1 non-overexpressing wild type mice were housed and sacrificed on day 30. Lung sections were subjected to immunofluorescence staining with confocal microscopy (LSM 510 META; Carl Zeiss Microsystems).
3. Western blot analysis and immunofluorescence staining
For western blot analysis, A549 cells were prepared by extracting proteins with RIPA buffer (50 mM Tris-HCl [pH 7.4], 1% [V/V] NP-40, 150 mM NaCl, and 1 mM ethylenediaminetetraacetic acid) supplemented with protease inhibitors (1 mM phenylmethylsulfonyl fluoride, 1 µM aprotinin, 1 µg/mL leupeptin, and 1 mM Na3VO4). Lung tissue was homogenized in RIPA buffer containing protease inhibitors. Equal amounts of proteins were resolved using 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred onto nitrocellulose membranes (Amersham Pharmacia Biotech Inc., Piscataway, NJ, USA). After blocking with non-fat dried milk, the membranes were incubated with primary antibodies for 1 hour at room temperature followed by detection using horseradish peroxidase conjugated secondary antibodies. Enhanced chemiluminescence detection was performed according to the manufacturer's instructions (Boehringer Mannheim, Mannheim, Germany). The relative abundance of protein was determined by quantitative densitometry using Image J software (NIH, Bethesda, MD, USA). All western blot densitometry data were normalized to β-actin.
For immunofluorescence analysis, mouse lung tissue was incubated at 4℃ overnight with anti E-cadherin (1:500, Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA) and N-cadherin antibodies (1:1,000, BD Bioscience, Bedford, MA, USA). Fluorescein isothiocyanate-conjugated donkey anti-goat antibodies (1:1,000, Santa Cruz Biotechnology Inc.) and goat anti-rabbit IgG-PE (1:1,000, Santa Cruz Biotechnology Inc.) were used as secondary antibodies for the localization of E-cadherin and N-cadherin in the mouse lung.
4. Antibodies
The antibodies used in the present study included anti-human E-cadherin (1:500, Santa Cruz Biotechnology Inc.), anti-human N-cadherin (1:1,000, BD Bioscience), anti-α-SMA (1:300, Abcam, Cambridge, UK), anti–β-actin (1:5,000, Sigma-Aldrich), anti-Smad3, anti-pSmad3 (1:1,000, Cell Signaling Technology, Danvers, MA, USA), anti-ERK1/2, anti-pERK1/2 (1:1,000, Cell Signaling Technology), anti-p38 mitogen-activated protein kinase (MAPK), anti-pp38 MAPK (1:1,000, Cell Signaling Technology), anti-TβRI (1:1,000, Santa Cruz Biotechnology Inc.), and anti-TβRII (1:500, Abcam).
Results
1. ApoA1 inhibits EMT in the TGF-β1–treated AEC and silica-induced lung fibrosis models
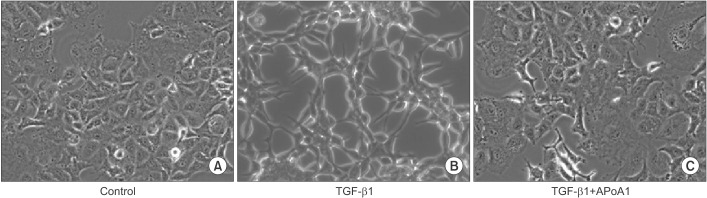
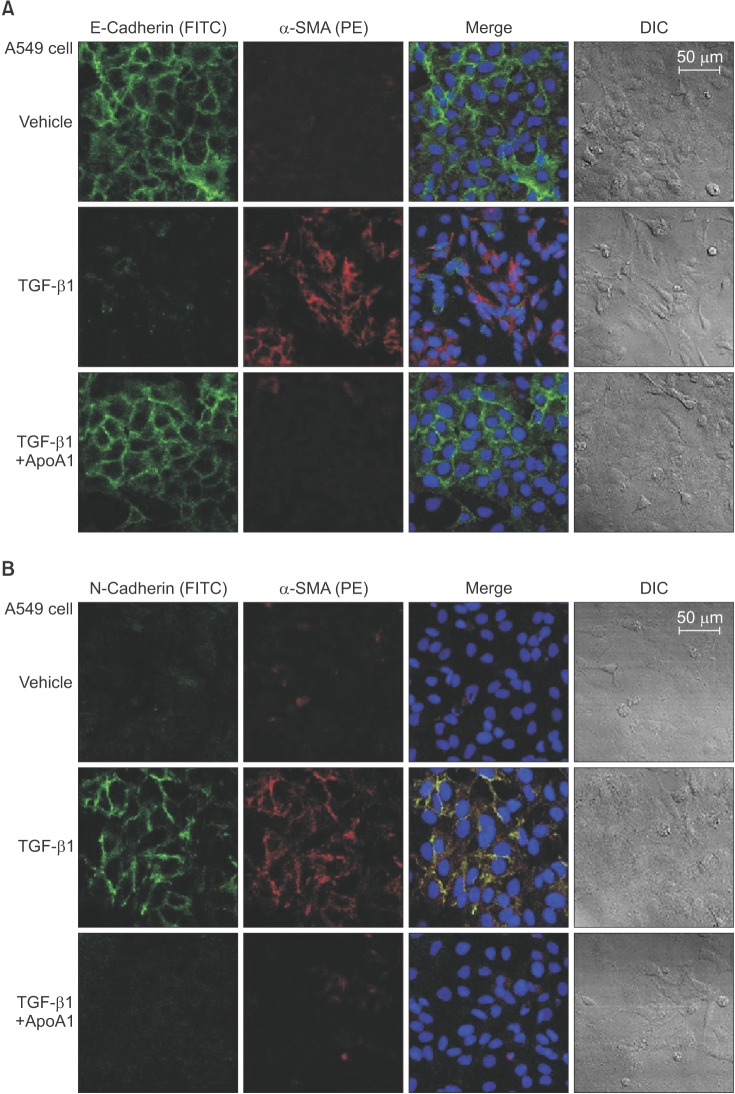
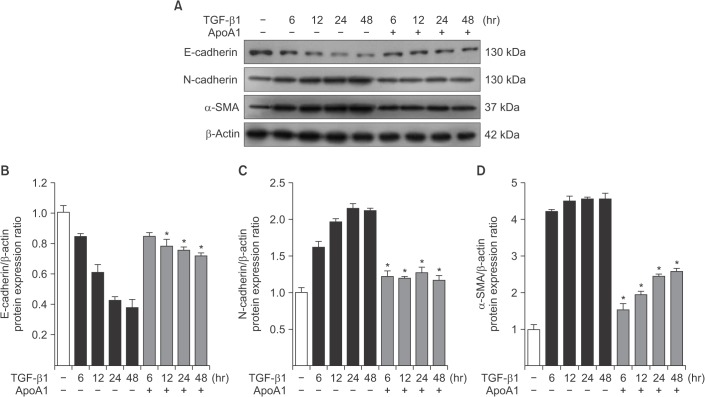
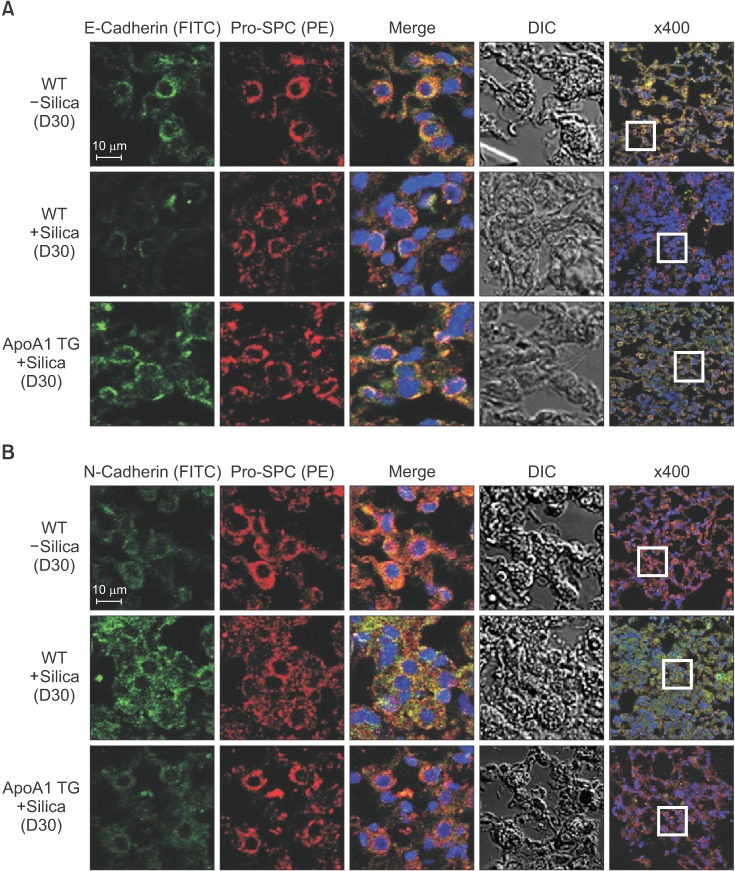
As previously reported, in response to TGF-β1, A549 cells were converted from an epithelial phenotype to a spindle-like mesenchymal phenotype, and treatment with ApoA1 restored the cells to the traditional polygonal epithelial shape (Figure 1). As expected, exposing the cells to TGF-β1 for 48 hours showed a reduced expression of the epithelial marker (E-cadherin) with increased expression of the mesenchymal marker (N-cadherin and α-SMA). These phenotypic changes were inhibited by treatment of ApoA1 (Figures 2, 3). Silica-treated mouse lung showed decreased E-cadherin and increased N-cadherin expression in the alveolar epithelium, shown as the pro-surfactant protein C, and overexpression of ApoA1 inhibited the changes in the epithelial and mesenchymal markers in the alveolar epithelium (Figure 4). These findings demonstrate that ApoA1 effectively inhibits both TGF-β1–induced EMT in the AEC and silica-induced EMT in vivo.
2. ApoA1 inhibits TGF-β1–induced Smad-dependent and non-Smad signaling pathways
We determined whether ApoA1 interferes with the TGF-β1 signaling pathways. We examined whether the inhibitory effects of ApoA1 against TGF-β1–induced EMT in A549 cells are mediated by changing the phosphorylated Smad2 or Smad3 expression, which have been demonstrated to be the major signal pathways in TGF-β1–induced EMT17.
Phosphorylation of Smad2 and Smad3 in A549 cells was markedly increased in a time-dependent manner by TGF-β1 treatment. When the cells were treated with ApoA1, phosphorylation of Smad2 and Smad3 were significantly reduced (Figure 5). This finding suggests that ApoA1 inhibits the Smad-dependent TGF-β1 signaling pathway. TGF-β1 also induced non-Smad responses, including ERK and p38 MAPK mediated signaling18. ApoA1 also reduced the phosphorylation of ERK and p38 MAPK induced by TGF-β1 (Figure 6). It appears that ApoA1 down-regulates both Smad-dependent and non-Smad signaling pathways induced by TGF-β1.
3. ApoA1 down-regulates TGF-β1 receptor expression
TGF-β1 signaling is initiated by the binding of TGF-β1 to TβRI and TβRII on the epithelial cell membrane19. Increases in the TβRI and TβRII protein expression levels were observed in A549 cells upon TGF-β1 treatment, whereas the addition of ApoA1 significantly reduced the expression of these proteins (Figure 7).
Discussion
In this study, we have shown that ApoA1 effectively inhibits TGF-β1–induced EMT, both in vitro and in vivo. ApoA1 remarkably reduced TGF-β1–induced expression of mesenchymal markers such as α-SMA and N-cadherin and recovered the expression of the epithelial marker E-cadherin in AECs (Figures 2, 3). We also observed that ApoA1 overexpression inhibits silica-induced EMT in the mouse model (Figure 4).
In the currently accepted paradigm, the main pathogenesis of IPF is thought to be recurrent AEC injury that leads to aberrant activation of AEC, creating the accumulation of collagen producing fibroblasts and myofibroblasts2021. Although the significance and relative contribution of EMT to the source of activated fibroblasts and myofibroblasts has not been definitely established, EMT has been increasingly proposed as one of the main mechanisms of excessive fibroblast proliferation in lung fibrosis2223. For the induction of EMT in vitro and in vivo, TGF-β1 is regarded as the prototypical cytokine24. TGF-β1–induced EMT starts with the activation of latent TGF-β1, which originally existed in the extracellular mileu, kept inactive by the latency-associated protein, and bound by the latent TGF binding protein. Then, TGF-β1 binds with the TβRII and TβRI, activating the receptor heterodimer and initiating the intracellular Smad and non-Smad signal pathways24.
TGF-β1 signaling occurs via both Smad dependent and non-dependent pathways25. The Smad dependent signal transduction system is the main process in the TGF-β1 signaling. When the extracellular TGF-β ligand binds to the cell membrane TGF-β receptor, the receptor-activated Smads (R-SMADS), Smad2 and 3, are phospholylated directly by the intracellular TβRI kinase domain, and then bind to the common mediator Smad (Co-SMAD), Smad4. These Smad proteins translocate into the nucleus, bind with Smad-binding elements as a transcription factor complex, and activate TGF-β1 target genes (e.g., α-SMA and collagen), as well as inhibiting epithelial genes (e.g., E-cadherin) through the co-association of various transcription factors1724. For the full TGF-β1 response, phosphoinositide 3-kinase/Akt or MAPK-targeted transcription factors bind to the critical response element of the TGF-β target genes in the Smad independent pathway. The ERK/MAPK cascade additionally phosphorylates R-SMADS, modifying Smad activity and enhances TGF-β1–mediated collagen I synthesis26. Besides such interaction with the Smad pathway in modulation and activation of transcription factors, non-Smad proteins are also involved in a variety of cellular responses, including cellular tight/adheren junction disassembly, and cytoskeletal rearrangement2425.
Each step in the TGF-β1–induced EMT signal pathway has been considered as a possible, effective therapeutic target for lung fibrosis27. Several studies to identify the proper therapeutic target in the TGF-β1–induced EMT signal pathway have been completed or are in progress2228293031. However, currently, there is no therapeutic agent that directly inhibits TGF-β1, due to its complex function and the expected side effects of direct TGF-β1 inhibition28.
In this study, treatment of ApoA1 attenuated TGF-β1–induced Smad2 and Smad3 phosphorylation in A549 cell (Figure 5). Moreover, ApoA1 also inhibited non-Smad signaling, particularly the ERK and MAPK pathways (Figure 6). Since both Smad-dependent and non-Smad pathways were inhibited by ApoA1 in our study, we evaluated further whether ApoA1 modulates TGF-β receptor expression. Rojas et al.32 reported that the TGF-β receptor levels regulate the specificity of the signaling pathway activation and the biologic effects of TGF-β1 and, in particular, that the TβRII expression levels are correlated with Smad signaling and MAPK-ERK signaling activation. We have shown that ApoA1 inhibits both TβRI and TβRII expression induced by TGF-β1 (Figure 7). The exact molecular mechanism by which ApoA1 modulates the expression of TGF-β receptors remains to be determined. We speculate that ApoA1 interferes with the binding or interaction between the TGF-β1 ligand and the TβRI-TβRII complex, resulting in down-regulation of downstream signaling molecules.
We examined the anti-EMT activity of ApoA1 through the murine model of silica-induced lung fibrosis using a previously reported method16. ApoA1 transgenic mice and a silica-induced lung fibrosis model were used because even distribution of ApoA1 in the alveolar epithelium and permanent chronic lung fibrosis are essential for EMT experiments in vivo. In our previous data, active TGF-β1 levels in the lung were significantly increased in the silica-treated mice16. In the present study, intratracheal treatment of silica decreased E-cadherin and increased N-cadherin expression in the alveolar epithelium. Overexpression of ApoA1 inhibits these EMT-related phenotypic marker changes. Taken together, we have demonstrated that ApoA1 blocks TGF-β1–induced EMT both in vitro and in vivo.
ApoA1 was originally known as the major apolipoprotein composed of high density lipoprotein cholesterol. In addition to cholesterol modulation, ApoA1 has been shown to possess anti-inflammatory activity3334. Recently, we reported that ApoA1 has a therapeutic potential on a bleomycin and silicainduced murine model of fibrosis, with possible mechanisms associated with anti-apoptosis, anti-oxidative activity and the ability to promote the generation of pro-resolutional mediators such as lipoxin A4151635. In addition to these results, these data support the idea that the inhibition of EMT by ApoA1 may at least partly contribute to its anti-fibrotic activity in the experimental lung fibrosis model. Further study is needed to investigate how ApoA1 modulates TGF-β receptor expression.
In conclusion, this study demonstrates that ApoA1 inhibits TGF-β1–induced EMT in AECs and a silica-induced lung fibrosis animal model. ApoA1 down-regulates Smad-dependent and non-Smad TGF-β1 signaling pathways and reduces TβRI and TβRII expression. These findings could be helpful in understanding the therapeutic effect of ApoA1 on lung fibrosis.
 XML Download
XML Download