

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Lung cancer is the leading cause of cancer-related mortality worldwide attributing 1 million deaths every year1. There has been no effective screening tool with survival benefit2,3, leading to search for novel, non-invasive method for early detection of lung cancers. Early detection includes the identification of lung cancer-specific biomarkers.
Pathogenesis of cancer is having been elucidated gradually as genetics and molecular biology evolves. Epigenetic alteration of genes with their rolls of tumor suppression is one of the central mechanisms of tumorigenesis other than genetic mutation. This promising field has been being researched for more than two decades, and methylation of the CpG islands in or near of promoter region of tumor suppressor genes is major composition of epigenetic mechanism. When methylation occurs, some other events such as deacetylations of histone residues readily accompany, and conformational change of DNA inhibits transcriptional factors and some proteins from binding for the transcription to be initiated. And loss of function of the tumor suppressor genes (TSGs), for example, is resulted without alterations of genetic sequences4-6. Epigenetic alterations are changes that could be reversed by demethylating agents such as azacitidine and some results from several studies on these agents with some kinds of cancers disclosed the effectiveness in survival rate and symptom relief, so application into clinical field was just started5.
In the present study authors hypothesized that if a gene could be a TSG by epigenetic mechanisms, more frequent methylation profiling of that gene should be observed in tumor tissue than adjacent non-tumor tissue in a subject, or in tissue from cancer-free control group. So we investigated methylation profiles of genes in nonsmall cell lung cancer (NSCLC) patients and compared with corresponding adjacent non-tumor lung tissues. We tried to search for non-small cell lung cancer-specific tumor suppressor genes of which their function is inhibited by epigenetic mechanism.
Materials and Methods
1. Patients & DNA extraction
Total of 10 patients with primary NSCLC who underwent lung surgeries in 2005 in Konyang university hospital were included. This study was approved by the Bioethics Committee of Konyang University Hospital and all of the participants gave written informed consent. All of the tumor and macroscopically normal lung tissue samples were obtained at the time of surgery, and were rapidly frozen in liquid nitrogen and stored at -80℃ until analysis. Also, normal lung tissue was obtained from two young healthy male patients with pneumothoraces through their bullectomies. Tissues samples were histologically confirmed by hematoxylin-eosin staining. DNA was extracted and prepared using the QiaAmp DNA mini kit® (Qiagen, Hilden, Germany).
2. Principle of the method
Methylation status of each promoter was detected by presence or absence of hybridization signals after PCR using samples digested with HpaII (methylation-sensitive enzyme) and MspI (Methylation-resistant enzyme). The hybridization signals on the Methyl-Scan DNA microarray are proportional to PCR status (Figure 1).
3. Microarray design
To make the microarray, we have chosen 50 genes (Apaf-1, ACCN1, APC, AR, BRCA1, CALCA, CALCR, CDH13, CDKN2A, CDKN2B, CFTR, COMT, DAPK1, EBR, EDN1, EPHA3, EPO, ESR1, FHIT, H19, hMLH1, HPSE, HTR1B, IL-8, JunB, LAMA5, LDHB, LRP2, LTB4R, MDR3. MGMT, MTHFR, MUC2, PGR, PIK3CG, PLS3, PTGS2, RAR-b, RB1, S100A2, SHP1, SKT11, SLC5A5, SMARCA3, SRBC, TFF1, TP73, TUSC3, VHL, and WT1) which have promoter regions for HpaII searched in NCBI database.
4. Digestion of DNA
The purified DNA (100 ng for each digestion) was digested with HpaII (20 units / 2 uL) and MspI (40 units / 2 uL), respectively, and then genomic DNA was re-digested with same enzymes to ensure complete digestion and reduce the background noise signals. Each digestion was followed by incubation at 37℃ for 2 hours and 15 hours, respectively. The second digested samples were purified with GeneClean Turbo kit® (Qbio, gene, Baton Rouge, LA, USA) according to the manufacturer's instructions. To ensure scanning, we used exogenous GAPDH cDNA as control.
5. Multiplex PCR and target labeling
Multiplex PCR amplification was done with un-digested and HpaII-, MspI-digested DNA with 9 primer sets to label 50 target promoter regions. During amplification step, fluorescent dyes were incorporated into the amplicons; Cy3-dUTP in MspI-digested targets, Cy5-dUTP in HpaII-digested targets, and undigested samples labeled with Cy5-dUTP by same multiplex PCR.
6. Hybridization and methylation DNA microarray analysis
After PCR amplification all the amplicons were mixed, microarray hybridization was performed at 65℃ for 4 hours in the humidified chamber (GenomicTree, Daejeon, South Korea). The hybridized microarray was imaged by scanner (Axon Instruments, CA, USA). If the signal intensity of HpaII amplicon is 2-fold greater than that of MspI amplicon, the target region was considered to be methylated, while less than 2-fold was considered to be unmethylated.
Results
1. Patient characteristics
There were 5 male and 5 female patients, the mean age was 62.9 years old. All of 5 male patients had smoking history with forty six of mean pack-years, two of them had squamous cell carcinoma, and the remainders had adenocarcinomas. 5 female patients were all non-smokers, 3 of them had adenocarcinomas, one had squamous cell carcinoma, and the remainder had other kind of NSCLC, respectively (Table 1).
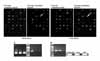
2. Results from microarray
The methylation microarray assay was performed for tumor tissue and adjacent normally-appearing tissue using HpaII-MspI methylation microarray. The PCR was successful for 21 genes (APC, AR, BRCA1, CALCA, CDH13, CFTR, EDN1, EPHA3, H19, HTR1B, LAMA5, LDHB, LTB4R, MTHFR, MUC2, PIK3CG, PTGS2, RAR-b, S100A2, SHP1, and SRBC) from 50 genes, others were not available because of PCR failure (Figure 2). APC, AR, RAR-b, HTR1B, EDHA3, CFTR and were hypermethylated in tumor tissue than adjacent normally appearing tissue. CDH13, CALCA, LAMA5, MTHFR, PIC3CG, S100A2, SHP1, EDN1, H19, MUC2, and LTB4R were commonly methylated in tumor and non-tumor sample and we found that these genes were hypermethylated in normal control, as well (Figure 3). Among the rest of genes excluding aforementioned eleven genes, four genes (SRBC, LDHB, BRCA1, and PTGS2) were not methylated in tumor tissue. Then six genes, APC, AR, RAR-b, HTR1B, EPHA3, and CFTR, finally turned out to be more frequently methylated in tumor tissue and unmethylated in non-tumor tissue and control group (Table 2).
3. Results from gel electrophoresis
The results for 22 genes including control (IFN) from four patients among subjects with gel electrophoresis were completely identical with those of DNA microarray (Figure 4).
Discussion
Aberrant promoter hypermethylation in the field of cancer has been researched vigorously for two decades. After hematologic malignancies with this mechanism and efficacy of administration of demethylating agents were studied, many other solid tumors are having been under investigation for years. Present studies are focusing largely on identification of TSGs functioning as DNA repair genes, cell cycle regulatory genes or chromosomal loci by epigenetic mechanism, and some of them more frequently methylated in specific cancers seem to be possible candidates7-10. While, hypermethylation profiles of several genes are being proposed as prognostic markers11-13.
With the recent development of large-throughput analysis technology, assays allowing CpG island methylation to be analyzed at a genome level were developed. Oligonucleotide-based methylation assays using PCR after bisulfite treatment were developed by Adorjan et al.14 and Shi et al.15 Methylation assays using DNA microarray, first introduced in 2001 by Yan et al.16, include a method comprising modifying the cytosine of genomic DNA into uracil, amplifying the modified DNA, polymerizing the amplification product into oligonucleotide or DNA-oligomer, and hybridizing the polymer in a DNA microarray17-19. To show the high accuracy of the method, we performed gel electrophoresis on IFN, which doesn't have a promoter region for HpaII, as a control. The results of gel electrophoresis were exactly identical with that of microarray. We suggest that microarray could be a promising method for mass screening in the future, with extended range of genes.
In the present study, CDH13, CALCA, LAMA5, MTHFR, PIC3CG, S100A2, SHP1, EDN1, H19, MUC2, and LTB4R were commonly in all three group; Among them CHD13, CALLA and EDN1 were described in previous studies to have hypermethylated promoter regions and relation with lung cancer; Takai et al. reported that DNA fragment B3 was hypermethylated in 14 of 20 (70%) primary cancers, and decreased EDN1 (endothelin-1) expression was observed in 16 of 20 (80%)20. In the meanwhile, in present study, EDN1 was hypermethylated in tumor and non-tumor tissue with equal frequency of 60%, and in all of the control tissue. Hypermethylation of the promoter of CHD13, a gene encoding H-cadherin, was frequently associated with silencing in a few cancers, including lung cancer21-23. Kim et al. reported the frequency of hypermethylation in tumor and non-tumor tissue is 26 of 88 tissue samples (29.5%) and 7 of 88 tissue samples (8%), respectively24. In present study, frequency of methylation of tumor and non-tumor tissue was 40%, and 30%, respectively with hypermethylation in all from control group. CALLA (NEP; CD10, EC 3.4.24.11) is a gene for cell surface endopeptidase inactivating the autocrine growth factors and inhibiting cell proliferation, and decreased activation of this gene have been found in lung cancers25-27. There could be some considerations for hypermethylation in normal tissue seen in present study; (1) the method with enzyme could have been too sensitive, that allowed false-positive results; (2) the expressions of genes are controlled by normal methylation process during the course of embryogenesis and development, as 'aging process', and the persistently functioning genes in adults are localized to specific tissue for specific functions28-30, that some of the genes above could be normally silenced by methylation in lung tissue.
Findings from our present study showed the 6 genes of APC, AR, CFTR, EPHA3, HTR1B, and RAR-b more frequently hypermethylated in tumor tissue than corresponding normal tissue or control, which suggests that their methylation may be a tumor-restricted event. And AR, APC and RAR-b are well-known to be TSGs silenced with epigenetic alteration for NSCLC from the previous studies7,31-35, that CFTR, EPHA3 and HTR1B are worthy to be studied further as possible candidates of TSGs by epigenetic mechanism for NSCLC.
Cystic fibrosis transmembrane regulator gene (CFTR) is known to be related to chronic idiopathic pancreatitis, a well known risk factor of pancreatic cancer and there were several studies to show that mutation of CFTR (Δ F508) would be associated with malignancies, including pancreatic cancer36,37. While, recently a few novel functions of CFTR has been reported other than as ion channels in airway epithelial cells in cystic fibrosis36-38; (1) As part of a multiprotein complex at the cell surface, CFTR might interfere with the expression of several gene products, and participate in the signaling pathways of inflammatory response conjunctly with NF-κB, mitogen-activated protein kinase/extracellular signal-regulated kinase (MAPK/ERK) and activated protein-1 (AP-1). And when AP-1 is acting associated with transcription co-factors which promote cell proliferation and inhibit apoptosis, this would result in respiratory epithelium carcinogenesis39. (2) it also permeate anti-oxidant-reduced glutathione (GSH) out of epithelial cells, resulting in the low level of intracellular glutathione, that leads to insufficient protection from oxidative stress such as smoking, and damage and apoptosis of cells occur. This mechanism is related with high cellular level of BAX which is a pro-apoptotic member of Bcl-2 family, and in cystic fibrosis which has CFTR mutation, the level of BAX would be lowered.
Recently, Brantley et al.40 reported that EPHA2 and EPHA3, which are in soluble form, are thought to have a role in inhibiting angiogenesis and tumor growth, and treatment with soluble EphA2-Fc or EphA3-Fc receptors inhibited tumor angiogenesis in cutaneous window assays, and tumor growth in vivo, in the report of the first functional evidence for Eph A class receptor regulation of pathogenic angiogenesis induced by tumors and support the function of EPHA receptors in tumor progression. As far as we know, there is no study undertaken on hypermethylation of promoter region of EPHA3 with decreased expression.
Takai et al.20 reported aberrant hypermethylation of CpG island in the promoter region of HTR1B (hydroxy-tryptamine receptor 1B) was seen in two squamous lung cancer cell lines, EBC-a and LK-2, while it was demethylated in normal human bronchial endothelium (NHBE). And when LK-w cells were treated with 5-aza-2-deoxycytidine, H6 fragment was demethylated and expression of HTR1B was restored. Maybe due to the heterogeneity of clinical samples of both lung cancer tissue and normal tissue, the results of methylation profile and expression of HTR1B was not exactly consistent with that with lung cancer cell lines. The role of HTR1B silencing in lung carcinogenesis is not clear but the authors suggested that it is of great interest to examine the role of serotonin and its signaling from its type B receptors.
Some limits in this study was the small number of patients and subject genes, but this study showed hypermethylation profiles of several genes that could be TSGs of lung cancer with microarray, which is not widely used in Korea yet, with high accuracy of the method.
DNA microarray has strengths of rapidity, mass analysis, easy to managing and also, high accuracy, suggesting the role of screening tool for cancers with DNA hypermethylation profiles as novel biologic markers. In the present study, CpG islands of 6 genes including APC, AR, CFTR, EPHA3, HTR1B, and RAR-b were hypermethylated in tumor tissue than tumor adjacent tissue or control group which was assessed with microarray. The results from the present studies and previous reports on CFTR, EPHA3 and HTR1B support that these genes possibly be novel candidates of TSGs in NSCLC. The functional study for each candidate gene is required for the confirmation of the genes as TSG by hypermethylation mechanism. Further study with this design with larger number of patients and whole human genes would be required.





 XML Download
XML Download