

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Vaccines have been developed for many infectious diseases since the early twentieth century and they have provided numerous benefits to human society. One of these benefits is the increased lifespan caused by the prevention of infectious diseases. However, the types of diseases have changed alongside the increased lifespan caused by the prevention of infectious disease. These new health threats are chronic diseases such as cancer, cardiovascular diseases, diabetes, and obesity [1]. Many therapeutic vaccine approaches have been developed for preventing chronic diseases [2]. In this review, we discuss the prophylaxes and therapeutic vaccinations that could be used to reduce the incidence of obesity and diabetes. In particular, we describe the possibility of treating pathogen-induced obesity.
Vaccine-Based Approaches for Treating Obesity and Diabetes
Obesity is classified as one of the eight significant causes of chronic disease by the World Health Organization (WHO). Sixty-five percent of the world's population lives in countries where overweight and obesity kills more people than underweight [3]. The number of health problems linked to chronic diseases is increasing dramatically and obesity is related to numerous chronic diseases. Many studies have also shown that obesity has a specific relationship with type 2 diabetes [4-6].
Obesity causes many problems in humans, but the treatment options for obesity are very limited. It is widely accepted that a combination of dieting and physical activity is the most effective way of reducing obesity [7]. However, these methods are highly dependent on the motivation of individual subjects. Thus, healthy practices are readily stopped and subjects may return to their previous weight, which is called the yo-yo effect. Therefore, several medications are being developed for treating obesity. However, current medications only have a beneficial effect if the treatment is sustained continuously [8]. In addition, bariatric surgery may lead to permanent weight loss but this method has significant perioperative risks and an economic burden for the patient [9,10]. To address the problems of obesity treatment, recent studies have shown that therapeutic vaccines may be new targets for the development of anti-obesity medications. The therapeutic vaccines could treat alternatively the obesity by suppressing the appetite-stimulating hormone and blocking absorption of nutrients.
One therapeutic strategy is to block endogenous signals to reduce the appetite and body weight. In particular, immunization against the gut peptide, ghrelin, has entered phase I and IIa trials as an anti-obesity therapy [11]. Ghrelin is produced mainly in the small and large intestines, although it is also secreted by the lungs, pancreatic islets, gonads, kidney, and brain [12]. Ghrelin is an appetite-regulating hormone and a contributor to mealtime hunger [13]. The levels of ghrelin increase before meals and decrease after feeding [13]. Ghrelin has a wide range of biological functions. In particular, it promotes weight gain by increasing the appetite and food intake, while reducing the energy expenditure [14,15]. However this vaccine could cause emotional side effects. To identify the possibility of its use as a vaccine, synthetic ghrelin analogs were conjugated to haptens [16]. Mice immunized with the ghrelin peptide sequence gained less body weight and had lower body fat and circulating leptin levels. In addition, a therapeutic vaccine has been tested that used a noninfectious virus to carry ghrelin to elicit an immune response to obstruct this hormone. This vaccine induced antibodies against ghrelin, showing that it can suppress this hormone. Vaccinated mice had higher levels of anti-ghrelin antibodies compared with unvaccinated mice. Moreover, the vaccinated mice had a lower food intake and increased energy expenditure, i.e., obese mice had 82% of the food intake consumed by the control mice after the first vaccination injection and only 50% of the food intake of the control mice after the final vaccination [16]. In addition, the vaccinated obese mice exhibited reduced neuropeptide Y (NPY) expression. NPY is the most potent signal that increases the appetite via the central nervous system [17,18].
Furthermore, a 'flab jab' vaccine has been reported that reduced the body weight by 10% four days after a single injection [19]. The vaccine was designed to block the effects of somatostatin, which has many different functions in various organs. In particular, somatostatin can suppress the release of pancreatic hormones. The release of somatostatin from the pancreas prevents insulin secretion and reduces the release of glucagon. Blocking this growth hormone has positive effects on obesity in obese animal models and in human studies. For example, vaccinated mice produced antibodies against somatostatin and their food intake was reduced compared with control mice at six weeks after vaccination. The vaccinated mice lost 12-13% of their body weight at four days after the first injection, although all of the mice consumed similar amounts of food [19]. Mice treated with the two vaccines, JH17 and JH18, exhibited 4% and 7% increases in their body weight, respectively, compared with their body weight at the start of the study. By contrast, control mice exhibited a 15% increase in their body weight. However, this study had several limitations, such as problems with the control vaccination, the overall weight gain, and the unrealistic volume of vaccine required [19]. In addition, this vaccine could cause steatorrhea, diarrhea, gastrointestinal cramps, and occasional nausea. Therefore, somatostatin vaccines can reduce weight gain and the final body weight compared with the baseline weight, although there are some problems with this approach.
Active vaccination against target molecules is another therapeutic vaccine approach. The recombinant virus-like particle (VLP) has a similar structure to viruses but lacks sufficient genetic information to replicate [20,21]. Thus, this type of vaccine is less dangerous compared with live virus vaccines. Moreover, VLPs derived from the coat protein of bacteriophage Qβ have no preexisting immunity [22]. During VLP assembly, single-stranded RNA triggers Toll-like receptors 7 and 8 [23], and these receptors induce co-stimulatory molecules and cytokines that promote antigen-specific IgG2a responses [24,25]. Chemical conjugation can be used to attach antigens to the surface of Qβ-VLP, thereby triggering potent antibody responses in animals and humans. In particular, VLP-based vaccines can induce antibodies against interleukin (IL)-1β to treat type 2 diabetes [26]. The cytokine IL-1β triggers islet cell apoptosis and is a primary agonist in the loss of beta cell mass in type 2 diabetes. Therefore, the prevention of IL-1β may improve the symptoms of type 2 diabetes patients [27,28]. When mice were immunized with the full-length IL-1β protein coupled to Qβ-VLP, they produced high neutralizing antibody titers with specificity for IL-1β [29]. The antibody response against the mutant form of IL-1β conferred protection in a rheumatoid arthritis animal model and the response also ameliorated the diabetic phenotype in a diet-induced obesity model [30].
Relationship between Obesity and Vaccination
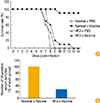
Obesity is a trigger of metabolic disease. Interestingly, the immune system is functionally linked to obesity. In particular, lymphoid tissues appear to be very sensitive to nutrient imbalances [31]. Therefore, these tissues have effects on metabolic pathways and the functions required for immune defense. Any impairment of the immune response may make obese individuals more susceptible to infection. In addition, it is well known that the effect to vaccination is reduced by obesity (Fig. 1) [32]. A previous study demonstrated an association between low antibody levels, obesity, and the response to hepatitis B vaccines [33]. Another study showed that a standard hepatitis B vaccine had a 71% protection rate in obese adults (body mass index >30) compared with 91% in the healthy control group [34]. A further study investigated the mechanism related to the reduced antibody response to vaccination [35], where ob/ob obese mice exhibited 21% and 12% lower levels of pre-B and immature B cells, respectively, compared with wild type C57BL/6 mice [35]. These results indicate that obesity can modify the immune system to produce an immunodeficient state, including altered lymphocyte and monocyte functions. Diet-induced and genetic obesity increase the susceptibility to bacterial and viral infections in humans and animals. Therefore, it is necessary to perform intensive studies to determine the effects of obesity on the efficacy of vaccinations.
Adenovirus 36 (Ad36) Vaccine as a Potential Anti-obesity Agent
Recently, many studies have reported that infections with pathogens can induce obesity and metabolic diseases. Thus, preventing and treating infection by selected pathogens may help to reduce the risk of the adverse effects of obesity.
Of the known obesity-inducing pathogens, human Ad36 induces adiposity but paradoxically it improves insulin sensitivity [36-38]. Ad36-infected animals have a higher body weight and fat content, but hypolipidemia compared with control animals. There is a strong association between Ad36 infection and human obesity [36-38]. Ad36 is a human adenovirus that belongs to serotype D and it is antigenically distinct [39]. According to epidemiological data, 30% of obese and 11% of non-obese adults in the United States possess antibodies against Ad36 [40]. Moreover, it has been reported that there is a relationship between viral infections with Ad36 and obesity in several nations [41-44]. However, research into the treatment of viral infections and vaccine development is still lacking in this area.
A previous study tested the possibility of using an inactivated Ad36 vaccine [45], where Ad36 was inactivated using ultraviolet (UV) irradiation. In trials, mice were inoculated with live Ad36, UV-inactivated Ad36, or the medium alone. Live Ad36 increased the size of the epididymal fat pad at 4 days post-inoculation, whereas UV-inactivated Ad36 did not cause an increase [45]. Furthermore, our group demonstrated a 'proof of concept' using an Ad36 vaccine (unpublished data). We injected mice with UV-inactivated Ad36, live Ad36 as the positive control, and medium as the negative control, before challenge with live Ad36 (unpublished data). The mice were sacrificed and their organs were used to study the chronic effects at 90 days post-challenge. Live Ad36-injected mice had larger epididymal fat pads and exhibited inflammation of their fat deposits. However, the body weight and epididymal fat levels of the UV-inactivated Ad36-injected mice were similar to those of the control mice. Moreover, treatment with UV-inactivated Ad36 decreased the level of inflammation in the fat deposits. Indeed, inactivated Ad36 reduced the levels of inflammatory cytokines and immune cells. Overall, these results show that UV-inactivated Ad36 can prevent increases in body fat and inflammation. Therefore, inactivated Ad36 may be a potential vaccine to prevent Ad36-induced obesity.
Conclusion
Hormones have been tested as targets to treat 'conventional' obesity by developing therapeutic vaccines. The incidence of pathogen-induced obesity is increasing, but the development of vaccines to treat these pathogens is insufficient at present. Several studies have demonstrated the 'proof of concept' of the potential application of a prophylactic vaccine against Ad36 as an anti-obesity agent. An inactivated Ad36 vaccine can prevent increases in the body weight and fat content, as well as reducing inflammation in fat deposits. However, this type of anti-obesity vaccine research is just beginning. Therefore, more intensive studies are required to develop prophylactic and therapeutic vaccines to prevent obesity.
 XML Download
XML Download