

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Morphine (C17H19O3N) is one of the strongest known anal gesic compounds [1]. The prevalence of opioid abuse is increasing globally, particularly in women of child-bearing age. Opioid administration during pregnancy can delay embryonic development, induce preterm labor, cause fetus death, produce chromosomal anomalies and neural tube defects, and result in low birth weight [2-7]. Infants of opioid-dependent mothers can display several behavioral abnormalities including hyperactivity, lower mental acuity, and motor development index [2, 3, 5].
An animal model has been used to document the neurotoxicity of high dosages of morphine [8]. Chronic administration of morphine can induce neuronal apoptosis in the central nervous system [9]. Morphine neurotoxicity was also shown in a human neuron cell culture model [10]. Morphine sulphate reduces the number of neurons and the thickness of cortex of cerebrum in mice neonates [11]. Morphine decreases the number and diameter of Purkinje cells, and the thickness of both the molecular and granular cerebellum layers of adult rats [12]. We previously reported that maternal morphine exposure during gestation reduces Purkinje cells in the one-day-old mice off spring [13].
These results highlight the concern regarding the high prevalence of opioid abuse in the world, particularly in women of child-bearing age, in light of the continued cerebellum development in the postnatal period. Very little is known of the effect of morphine sulphate on cerebellar development during infancy and after weaning. The present study was conducted to clarify the Purkinje cell and cortical layer alterations in the cerebellum of mice pups born to morphine-exposed mothers.
Materials and Methods
This experimental study was performed at the Gorgan Faculty of Medicine, Golestan University of Medical Sciences, Gorgan, Iran. Guidelines on the care and use of laboratory animals and approval of the ethic committee of Golestan University of Medical Sciences were observed.
Experimental animals
Balb/c mice, weighing 28-30 g, and 8-9-weeks-of-age were maintained in a climate-controlled room under a 12-hour alternating light/dark cycle at a controlled temperature of 20-22℃. Dry food pellets and water were provided ad libitum.
Drug
Each vial contained l ml of morphine sulphate (Darou Pakhsh, Tehran, Iran) dissolved in 3.3 ml of sterile 0.9% saline. The morphine sulphate was intraperitoneally (i.p.) injected into each mouse at a concentration of 10 mg/kg body weight.
Treatment regimen
After 2 weeks of acclimation to the diet and the environment, females were randomly divided into control and treated groups. Twelve female mice in the treated groups received morphine sulphate as detailed above during 7 days before mating and during the gestational period (GD 0-21), at 18 days after delivery in experimental group I and 32 days after delivery in experimental group II. Twelve female mice in the control groups received an equivalent volume of normal saline i.p. during the 7 days before mating, and at GD 18 (group III) or 32 (group IV). After parturition, on postnatal day 18 (P18) and 32 (P32), six mice pups of each control and experimental group were randomly selected and scarified quickly with chloroform anesthesia. The brain was exposed and fixed by immersion into 10% neutral-buffered formalin. After processing, 6 µm-thick sections were acquired using a model HM 325 microtome (Microm, Walldorf, Germany). Serial coronal sections were selected according to anatomical landmarks corresponding to bregma (-6.255 mm to -6.955 mm) of the cerebellar simple lobule. The sections were stained with cresyl violet for morphometric examination.
Morphometric techniques
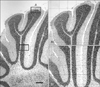
In each sample, six similar sections of simple lobule of cerebellum were selected and images of six separate fields in the apex and depth position were captured using a model BX 51 microscope and DP12 digital camera, with associated OLYSIA autobioreport software (Olympus Optical, Tokyo, Japan). Morphometric analyses of cerebellar cortex including thickness of the molecular layer (ML), Purkinje cell layer (PCL), and granular cell layer (GCL) were measured by low magnification (40× objective) (Figs. 1-3). Morphometric analyses of the Purkinje cells, which included the linear density of Purkinje cells, soma area, perimeter and diameter of Purkinje cells in the apex and depth of folium, were measured at high magnification (100× objective) (Figs. 1-3). Purkinje cells that exhibited an appropriate grid was applied on the image and the linear density of the cells was determined from the number of cells per 1,000 µm length of Purkinje cell line. Also the relative area, perimeter and diameter of the cell body was calculated by the software only for those Purkinje cells with a fixed geometric point of nucleus and with prominent nucleolus (Figs. 2, 3). The results are expressed as the mean of thickness of layers, diameter (µm), soma area (µm2), and linear densities of the Purkinje cells in each group.
Results
Weight of mice
No significant increase in average body weight, brain weight, and brain-to-body weight ratio were found between the morphine sulphate and control groups at P18 and P32 (Table 1).
Thickness of cerebellar cortex layers
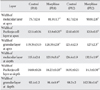
There was significant increase in the width of ML and granular layer at apex and depth of the simple lobule in the treated mice in comparison with controls at P18 and P32. The thickness of the Purkinje layer of cerebellar cortex was significantly reduced at P18 and P32 in the morphine-exposed mice in comparison with the controls (P<0.05) (Table 2, Fig. 2).
Quantitative characteristics of the Purkinje cells
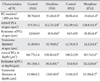
Concerning linear Purkinje cells density, the number of Purkinje cells per 1,000 µm length of Purkinje cell line of the experimental groups at P18 (23.40±0.5) and P32 (23.45±1.4) was reduced in comparison with control groups (28.70±0.9 at P18, 28.95±0.4 at P32) (both P<0.001) (Fig. 3). Concerning the area, perimeter, and diameter of the Purkinje cells, the cell area and perimeter in apex and depth of simple lobule in P18 and P32 cerebellar mice in the treated groups were significantly reduced in comparison with mice in the control groups (Fig. 3). The transverse diameter of Purkinje cells in the experimental groups was reduced in comparison with the control groups at P18 and P32 (P<0.001) (Table 3).
Discussion
According to our findings, administration of morphine sulfate before and during pregnancy, and during lactation, causes Purkinje cell loss and reduces cell size in 18- and 32-day-old infant mice. The toxic effect of morphine continues on the Purkinje cells of cerebellum even after the lactation period.
We previously reported that the maternal morphine exposure during gestation reduces Purkinje cells in one-day-old mice offspring [13]. Furthermore, two studies reported that morphine sulfate administration can cause neurotoxic effects on the cerebellum and cerebral cortex [11, 12]. One of these studies showed that oral morphine exposure during the GD reduces both cortical thickness and the number of neurons in the developing fetal frontal cerebral cortex [11]. The second study reported that the long-term administration of morphine sulphate in adult rats significantly reduced the Purkinje cells [12]. Other substances, such as alcohol and nicotine, can cause cerebellar Purkinje cell loss [14, 15].
Regarding the effect of morphine on the central nervous system, several possible mechanisms can be considered. Loss of the Purkinje cells in morphine treated animals can be due to apoptosis and or necrosis [16]. On the other hand, morphological alterations of astrocytes due to morphine can increase both Ca2+ and production of carbonyl oxidation, which subsequently promote apoptosis and or necrosis in neurons [17]. Furthermore, morphine can block the proliferation of neuroblasts in ML by preventing of DNA synthesis [18]. Several studies have shown that acute opioids exposure blocks the proliferation, differentiation, and survival of neuroblasts and astroglia of the cerebellum [19-23]. Reduction of the Purkinje cells in morphine chronic users has been reported [24]. The involvement of opioids in cerebellar growth regulation has been revealed by experimentally perturbing the endogenous opioid system. Endogenous opioid peptides and receptors are widely expressed by developing cerebellar cells [25-29]. Although heroin and morphine preferentially activate µ opioid receptors, at high concentrations they can activate δ and κ receptors [30]. Continuous opioid receptor blockage accelerates cerebellar growth in postnatal rats, while over-stimulating opioid receptors, as occurs with opiate drugs, retards cerebellar growth [19, 23, 31]. Also, several studies have reported that morphine as heroin can reduce Purkinje cell proliferation [16, 32], cell differentiation, and increase Purkinje cells death in the cerebellum [21, 28]. Neurotoxic effects of opioids can be inducing by the N-methyl-D-aspartate receptor-caspase pathway [8]. Indeed, cell death can be due to mitochondrial damage [33-35].
Another possible mechanism for the alteration of the Purkinje cells may be the significant decrease of calbindin protein as a neuroprotective agent in neurons [36, 37]. Furthermore, opioids may block neuronal activity, causing the neurons to receive internal signals to commit suicide (apoptosis) [38]. Other possible mechanisms for Purkinje cells loss may be due to derangement of the Purkinje cells in the rat cerebellum following prenatal exposure to morphine. Reelin has been shown to play an important role in the migration of neural cells, prompting the suggestion that the decrease in Reelin by X-ray irradiation is an important factor for the derangement of Purkinje cells [39].
Also, morphine presently induced a significant reduction of the thickness of the Purkinje layer. Our finding is similar to other studies [12, 40]. The cause of the reduction of the thickness of Purkinje layer may reflect decreased Purkinje cell size. Morphine presently induced a significant increase in the thickness of the molecular and granular layers in all treated mice. Our finding contrasts with other studies [12, 41]. Bekheet et al. [12] reported that morphine significantly decreased the thickness of molecular and granular layers in all treated animals. Also, another study reported that morphine induced a significant decrease in the molecular and granular layers thickness of the rat cerebellum [41]. The differences between the present and previopus findings regarding the thickness of molecular and granular layers of the cerebellum may be due to the timing of the studies. Our study was done in infancy but the other studies were done on adult animals. According to Simat et al. [42], proliferation and migration of basket and stellate cells in the ML is initiated after 7 postnatal days. Further studies are required to survey the type of cells and layers of cerebellum in different phase of postnatal periods.
In conclusion, this study reveals that morphine administration before and during gestational and lactation period causes Purkinje cell loss and reduction of the size and thickness of PCL of the cerebellar cortex in 18- and 32-day-old infant mice. Comparing the present findings with our previous study, we can conclude that the neurotoxic effect of morphine will be continued, even with discontinuation of exposure.



 XML Download
XML Download