

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The role of dietary long-chain polyunsaturated fatty acids (LCPUFA) in bone has been investigated in both healthy and osteoporotic states in a variety of species and developmental stages [1,2,3,4]. During growth, addition of ω-3 LCPUFA (eicosapentaenoic acid EPA and docosahexaenoic acid DHA) enhances the bone formation rate, the mechanism of which has largely been attributed to reduced synthesis of prostaglandin E2 (PGE2) in bone [2]. PGE2 is a potent stimulator of both bone formation and bone resorption [5]. The effect of PGE2 on bone is based on its amount in bone, which follows a concentration dependent pattern [2], with moderate concentrations encouraging bone formation, and high concentrations causing elevated bone resorption [6]. The ω-3 LCPUFA are precursors to PGE3, which is equally as potent as PGE2, in bone. However, conversion is less effective than for PGE2 from ω-6 LCPUFA (arachidonic acid ARA), resulting in a decrease in total PGE2. Reduction in the concentration of PGE2 may result in reduction of bone resorption [7], and enhancement of bone formation [2]. The relationships among dietary fat, calcium metabolism, and bone development are still not clear. Dietary lipids, depending on the type and amount ingested, may impair bone growth [8] or have no effect [3,9,10]. In addition, dietary lipids modify the fatty acid composition of cartilage and bone. Supplementation of the diet with fish oil resulted in elevation of EPA and DHA fatty acids in phospholipids of bone, while decreasing the concentration of ARA fatty acid compared to supplementation with corn oil or soy bean oil. Two main mechanisms have been suggested for the effect of alteration of specific dietary ω-6 and ω-3 LCPUFA ratios on bone; altered bone metabolism (formation or resorption), including osteoblast differentiation, modulation of PGE2 production, and inflammatory processes [2,11,12], and/or changes in Ca metabolism (absorption or excretion) [13,14], however, the exact effects of LCPUFA on Ca and bone metabolism remain unclear [9]. Alteration in the dietary ratio of w-6/w-3 and supplementation with specific LCPUFA (ARA, EPA, and DHA) in the diet is known to increase Ca transport across the mucosal membrane [15], Ca absorption [1], bone Ca content [1,3], and bone mass. On the other hand, decrease in Ca excretion [16] and bone loss were reported in later stage of life [17]. However, the amounts of specific ω-6 and ω-3 LCPUFA in the diet or in bone required for optimizing bone mass and mineralization or for slowing bone loss are not known [9]. The purpose of this study was to investigate different long term effects of consumption of dietary oil sources with varying omega-6/omega-3 (ω-6/ω-3) polyunsaturated fatty acids (PUFAs) ratios on the bone marrow fatty acids level, ex vivo prostaglandin E2 (PGE2) release, and mineral content of bone in rabbits, and to determine the role of gender to respond similarly or differently to this variation, and finally, to determine correlation between bone marrow ω-6/ω-3 ratio and mineral content in bone.
MATERIALS AND METHODS
Diets and Animals
Basal diet was purchased from the Arabian Agricultural Services Company (ARASCO), Riyadh, Saudi Arabia, and was prepared in accordance with its specification for rabbit feed (47152-Rabbit 18/14 Pellet, without fat). Five oils, namely soy bean oil (SBO), sesame oil (SO), fish oil (FO) (DHA 40%+ EPA 30%), and two types of marine brown microalgae oils of the genus Crypthecodinium cohnii; DHA 40% (40 g/100 g of fatty acids) and ARA 40% (40 g/100 g of fatty acids) were used in this study. The experimental diets were prepared by addition of the oil blend to the basal diet (70 g/kg diet), as follows: SBO diet, 70 g soy bean oil/kg diet; SO diet, 50 g sesame oil + 20 g soy bean oil; FO diet, 50 g fish oil + 20 g soy bean oil; DHA diet, 50 g DHA oil + 20 g soy bean oil; DHA/ARA diet, 25 g DHA oil + 25 g ARA oil + 20 g soy bean oil. Fresh diets were mixed weekly in order to avoid oil oxidation and kept refrigerated at 4℃ until fed.
A total of 45 weanling New Zealand white rabbits (male = 25, female = 20, 6-wk-old, weighing 500-1000 g) were obtained from the Experimental Animal Care and Experimental Surgery Center at the Faculty of Medicine, King Saud University, Riyadh, Saudi Arabia, and randomly divided by weight into five groups, where the weight difference between the members of each group did not exceed ± 100 g. All rabbits were housed individually in stainless steel cages under controlled temperature (25 ± 2℃) and relative humidity (50 ± 5%), with a 12-h light/dark cycle. Food and tap water were offered ad libitum throughout the experimental period (100 days). This study was approved by the Animal Ethics Committee of the College of Science, King Saud University (Ethic.RSC#5, 2nd, 2012).
Experimental diet
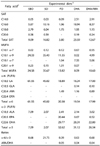
Fatty acid analysis of the formulated dietary treatments showed values of 0.39 to 21.75 for the ratio of ω-6/ω-3 fatty acids (Table 1). The lowest value was observed for the fish oil diet (FO) and the highest value was observed for the sesame oil diet. The amounts of linoleic acid LA (18:2, ω-6) and α-linolenic acid ALA (18:3, ω-3) ranged from 17 to 61.55 g and 2.07 to 7.09 g/100 g of the total fatty acids, respectively. The soy bean oil diet (SBO) contained the highest values of LA and ALA. The contents of ARA, EPA, and DHA ranged from 0.89 to 1.49 g, 0.52 to 20.44 g, and 22.8 to 29.77 g/100 g of the total fatty acids, respectively. The SBO and SO diets were devoid of ω-6 (ARA) and ω-3 (EPA and DHA) LCPUFA.
Sample collection
At the end of the experiment, rabbits were deprived of food overnight. They were then sacrificed immediately according to the procedures of the Experimental Animal Care and Experimental Surgery Center at the Faculty of Medicine, King Saud University, Riyadh, Saudi Arabia. Bones (tibia and femur) were obtained and bone marrow was removed as described in a previous study [18]. Bone samples were rinsed with normal saline (0.9% NaCl), dried using a lint-free paper towel, and stored in a plastic tube at -20℃ for one month until used for tibia PGE2 and femur mineral analysis.
Assays
Bone marrow samples were homogenized at 4℃ (Bench top Homogenizer 300 DS PRO Scientific, Inc., Oxford, CT, USA). Total lipids from 0.8 g of the homogenized tissue were extracted [19]. Extracted lipids and oils blend samples were transmethylated, using 14% boron trifloride in methanol, to fatty acid methyl esters (FAMEs) [20]. FAMEs were separated by gas chromatography and fatty acid concentration was expressed as weight percentage of each fatty acid in total fatty acids.
Bone organ culture was performed as described in the literature [2,9]. PGE2 analysis was performed in duplicate using a competitive enzyme-linked immunosorbent assay (ELISA) technique using a rabbit polyclonal antibody PGE2 kit (Oxford Biomedical Research, Inc., Oxford, MI, USA). PGE2 levels were expressed as ng/g bone.
Measurements of Ca, Mg, and Zn concentrations were performed in triplicate using an inductively coupled plasma mass spectrometer (ICP-MS 7500A, Agilent Technologies, Inc., Santa Clara, CA, USA), and P concentration was measured in triplicate using the molybdo-vanadate colorimetric method according to the AOAC [21]; optical density was measured at 410 nm using a UV/Visible 160A Shimadzu spectrophotometer, Kyoto, Japan. Mineral concentration was expressed as mg/g bone.
Statistical analysis
Data analysis was performed using SAS statistical software (version 9.2, SAS Institute Cary, NC, USA) and expressed as mean. Analyses of differences between male and female were performing by t-test at a significance level of P ≤ 0.05. Analyses of differences among the dietary treatment groups were performing by one-way ANOVA at a significance level of P ≤ 0.05; if significant differences were found, a Post-hoc analysis was performed using Duncan's multiple range test. Simple regression analysis was performed using Pearson's correlation coefficient for detection of correlations between bone marrow ω-6/ω-3 ratio, ARA/EPA ratio, ex vivo PGE2 release, and mineral content in bone.
RESULTS
Bone marrow fatty acids of rabbits
The bone marrow fatty acid profile of male and female rabbits was significantly influenced by and reflected the dietary lipid treatments (Table 2). In addition, significant differences in fatty acid profile were observed among the dietary treatment groups in both genders. Compared to the control (SBO) group, the lowest values of ARA and total ω-3 PUFA were observed in rabbits fed the SO diet in both males and females. Rabbits fed the FO diet showed the highest values of EPA, and total ω-3 PUFA in males, and EPA, DHA, and total ω-3 PUFA in females. Rabbits fed the DHA diet showed the highest values of DHA in female, but lowest values of LA, ALA, and total ω-6 PUFA in both males and females. Rabbits fed the DHA/ARA diet showed the highest values of 18:3 ω-6 and ARA, but the lowest values of EPA and DHA in both genders. However, the highest values of LA, ALA, and total ω-6 PUFA were observed in rabbits fed the control diet in both genders. In addition, significant differences in fatty acid ratios were observed among the dietary treatment groups in both genders (Table 2). Compared to the control (SBO) group, rabbits fed the SO diet showed the highest ω-6/ω-3 ratio (P ≤ 0.05); those fed the FO diet showed the lowest ω-6/ω-3 and ARA/EPA ratios (P ≤ 0.05); and those fed the DHA/ARA diet showed the highest ARA/EPA ratio (P ≤ 0.05).
Significant differences (P ≤0.05) in EPA and total ω-3 concentrations were observed between males and females. Significantly higher concentrations were observed for females fed the FO diet (P ≤ 0.05) than for males (Table 2). In addition, significant differences in ω-6/ω-3 ratio were observed between males and females (P ≤ 0.05). A significantly higher ratio was observed for males fed the DHA diet (P ≤ 0.05) than for females (Table 2).
Mineral content and ex vivo PGE2 level in rabbit bones
Mineral content and ex vivo PGE2 level of male and female rabbits are shown in Table 3. By the end of the feeding trial, a significant main effect of dietary treatment on femur Ca, P, Mg, and Zn contents was observed in both genders. Significantly higher Ca content was observed in males and females in the DHA/ARA, DHA, and FO groups (271.06 ± 6.97, 291.28 ± 4.56; 255.19 ± 5.16, 261.49 ± 2.26; and 244.66 ± 5.49, 249.15 ± 1.14 mg/g bone, respectively) compared to the control and SO groups (185.06 ± 2.41, 196.65 ± 2.08 and 213.55 ± 3.66, 174.03 ± 3.5 mg/g bone, respectively). Similarly, significantly higher Mg content was observed in males and females in the DHA/ARA, DHA, and FO groups (4.35 ± 0.13, 4.62 ± 0.17; 3.85 ± 0.14, 4.0 ± 0.04 and 3. 80 ± 0.27, 3.93 ± 0.2 mg/g bone, respectively) compared to the control and SO groups (2.95 ± 0.15, 3.17 ± 0.24 and 3.24 ± 0.1, 2.64 ± 0.04 mg/g bone, respectively).
Significantly higher P content was observed in males and females in the FO, DHA/ARA, SO, and DHA groups (136.12 ± 2, 136.42 ± 0.09; 134.24 ± 1.57, 134.51 ± 2.69; 132.19 ± 2.70, 135.54 ± 3.4; and 131.99 ± 2.46, 130.25 ± 1.30 mg/g bone, respectively) than in the control group (115.80 ± 5.18 and 122.79 ± 1.14 mg/g bone). In addition, significantly higher Zn content was observed in males in the DHA/ARA and the SO groups (0.10 ± 0.009 and 0.10 ± 0.008 mg/g bone, respectively) compared to the control group (0.08 ± 0.004 mg/g bone); and intermediate values (0.07 ± 0.005 and 0.07 ± 0.004 mg/g bone, respectively) were observed for the FO and the DHA groups. In females, significantly lower Zn content was observed in the FO group (0.07 ± 0.003 mg/g bone) and was slightly increased in the DHA/ARA group (0.10 ± 0.002 mg/g bone) compared to the control group (0.09 ± 0.004 mg/g bone); and intermediate values (0.08 ± 0.003 and 0.08 ± 0.01 mg/g bone, respectively) were observed for the SO and the DHA groups.
In addition, significant differences in Ca, Mg, and Zn, but not P content, were observed between males and females. Significantly higher Ca, Mg, and Zn content was observed in males fed the SO diet than in females, whereas significantly higher Ca content was observed in females fed the SBO diet than in males; however, no significant differences were observed between males and females in the other groups. Significantly lower ex vivo PGE2 release from tibia was observed in male rabbits fed the FO or DHA diets (6.88 ± 0.67 and 9.39 ± 0.52 ng/g bone, respectively) compared to the other groups (SBO, 12.59 ± 0.38; SO, 13.35 ± 1.1; and DHA/ARA, 12.31 ± 0.38 ng/g bone), while in females, no significant differences were observed among the dietary treatment groups (SBO, 11.52 ± 1.79; SO, 11.58 ± 0.79; FO, 9.95 ± 1.87; DHA, 10.58 ± 0.99; and DHA/ARA, 12.92 ± 0.96 ng/g bone). In addition, no significant differences in the ex vivo PGE2 level were observed between males and females in all groups.
Regression analysis
Regression analysis was performed to determine the correlations between bone marrow ω-6/ω-3 ratio, ex vivo PGE2 release, and mineral content in bone; a positive correlation was observed between ω-6/ω-3 ratio and PGE2 level (Fig. 1, panel A), a negative correlation was observed between ω-6/ω-3 ratio and Ca (Fig. 1, panel B), a negative correlation was observed between ω-6/ω-3 ratio and P (Fig. 1, panel C), a negative correlation was observed between ω-6/ω-3 ratio and Mg (Fig. 1, panel D), and a positive correlation was observed between ω-6/ω-3 ratio and Zn (Fig. 1, panel E).
In addition, correlations between bone marrow ARA/EPA ratio, ex vivo PGE2 release, and mineral content in bone indicated important associations.
A positive correlation was observed between ARA/EPA ratio and PGE2 level (Fig. 2, panel A), a positive correlation (P ≤ 0.05) was observed between ARA/EPA ratio and Ca (Fig. 2, panel B), a positive correlation was observed between ARA/EPA ratio and P (Fig. 2, panel C), a positive correlation was observed between (P ≤ 0.05) ARA/EPA ratio and Mg (Fig. 2, panel D), and a positive correlation (P ≤ 0.05) was observed between ARA/EPA ratio and Zn (Fig. 2, panel E). However, the correlation between ex vivo PGE2 release and mineral content in bone revealed a negative correlation between PGE2 level and Ca, P, and Mg (Fig. 3, panels A, B, and C, respectively), and a positive correlation between PGE2 level and Zn (Fig. 3, panel D).
DISCUSSION
This is the first report on the long-term effect of different dietary oil sources with varying ω-6/ω-3 ratios on bone marrow fatty acid profile, ex vivo PGE2 release from bone, and bone mineral content, including Ca, P, Mg, and Zn in male and female rabbits during growth.
There was a main effect of different dietary ω-6/ω-3 ratios on bone marrow EPA, total ω-3, ω-6/ω-3, and ARA/EPA levels. As the dietary ratio of ω-6/ω-3 declined from 21.8 to 0.4, the concentrations of EPA and total ω-3 rose; and, as the dietary ω-6/ω-3 ratio declined, so did the ω-6/ω-3 and ARA/EPA ratios. This study confirmed that different dietary oil sources varying in their ω-6/ω-3 ratios caused significant alteration of the fatty acid profile of bone marrow. Rabbits fed the FO diet (higher in ω-3 PUFA and lower in ω-6/ω-3 ratio) maintained a higher concentration of total ω-3 PUFA and lower ω-6/ω-3 ratio compared to those fed the SBO control diet with a higher ω-6 PUFA content. Those fed the SO diet (higher in ω-6/ω-3 ratio) maintained a higher ω-6/ω-3 ratio compared to those fed the control SBO diet. In addition, results of this study showed that bone marrow fatty acid profile in different groups reflected the effects of different dietary treatments [2,22,23].
The effects of varying dietary ω-6/ω-3 fatty acid ratios on bone marrow reflected the dietary level of ω-6 and ω-3 fatty acids fed to rabbits. As the dietary level of EPA and DHA rose, the concentration of ARA declined in bone marrow. The concentrations of EPA and DHA in bone marrow showed negative correlation (r = -0.52 and r = -0.88, respectively) with dietary ratio of ω-6/ω-3 fatty acids. The ω-6 and ω-3 fatty acids compete for the same desaturase and elongase enzymes with the substrate preference of ω-3 over ω-6 [24]. In addition, ω-6 and ω-3 fatty acids compete for incorporation into membrane phospholipids [25]. Thus, the result of the current study confirms the existence of competition between ω-6 and ω-3.
The levels of EPA and total ω-3 PUFA differed significantly between male and female rabbits. Significantly higher concentrations were observed in females compared to males. This finding suggests that the two genders incorporate ω-3 PUFA into bone marrow at a different rate and/or through a different mechanism. It has been suggested that females tend to have higher plasma levels and synthesis of ω-3 PUFA than males [26]. The finding of the current study contradicts other findings in the literature [27], where higher levels of EPA and total ω-3 PUFA in the femur were observed in male mice than in female mice.
The higher concentration of ω-3 PUFA found in bone marrow showed an association with reduced ex vivo PGE2 release in bone. In addition, as the dietary ratio of ω-6/ω-3 fatty acids approached 0.4, PGE2 showed a significant decrease in male and female rabbits fed the FO diet compared with other groups. This finding is in agreement with results reported in previous studies [2,4,9], where similar response of ex vivo PGE2 release was found in bone of rats. PGE2 is an important factor in regulation of local bone metabolism, including both bone resorption and bone formation [28]; in particular, it is a potent inducer of bone resorption at high concentrations [29]; while at low levels, it is a stimulator of bone formation [30]. Therefore, the ω-3 PUFA might have lowered the rate of bone resorption as a consequence, at least in part, of their suppressive effect on PGE2 biosynthesis [4]. The change observed in PGE2 with the lower dietary ratio of ω-6/ω-3 fatty acids reflects more favorable condition for promotion of bone formation and reduction of bone resorption, thus improving bone mass during growth.
Significantly higher Ca, P, and Mg contents were observed in male and female rabbits fed the FO diet low in ω-6/ω-3 ratio than in those fed the control diet with high ω-6/ω-3 ratio, despite their low intake of these minerals as a consequence of low food intake. In the current study, the reduction in ω-6/ω-3 ratio through inclusion of ω-3 LCPUFA in the diet from fish oil was shown to increase mineral content in bone of growing rabbits. This result is not in agreement with those of previous studies, where no such effects were observed in growing rats [4,9,10], while it is in agreement with another study [3], which found that inclusion of ω-3 LCPUFA in the diet from fish oil resulted in a significant increase in Ca and P contents in the bones of quails. This discrepancy might be related to short duration of these studies, 15 - 63 days versus 100 days for the current study and 210 days for another study [3]. Despite conflicting results, the current study does support a beneficial role of fish oil in bone Ca, P, and Mg contents.
Significantly higher Ca, P, and Mg contents were observed in male and female rabbits fed DHA or DHA/ARA diets low in ω-6/ω-3 ratio than in those fed control diet with high ω-6/ω-3 ratio, despite their low intake of these minerals as a consequence of low food intake. Of particular interest, male and female rabbits fed DHA or DHA/ARA diets maintained a Ca/P ratio of 2:1 (1.94 and 2; 2 and 2.2, respectively) compared to the other groups. In the current study, the reduction in ω-6/ω-3 ratio through inclusion of ω-3 LCPUFA in the diet from marine algae oils was shown to increase mineral content and maintain optimal Ca/P ratio in bone of growing rabbits. Therefore, it is possible that these novel oils may be promising dietary sources for promotion of bone mineralization during the growing stage of rabbits.
In contrast, significantly lower Ca, P, and Mg contents were observed in male and female rabbits fed the control diet; rich in ω-6 PUFA, with high dietary ω-6/ω-3 ratio than in those fed the DHA, DHA/ARA, or FO diets with low dietary ω-6/ω-3 ratio. This result clearly demonstrates that the elevation in ω-6/ω-3 ratio had a detrimental effect on bone mineralization, thereby bone growth.
Regarding femur Zn content, a significant increase in the concentration of the low ω-6/ω-3 ratio found in the DHA/ARA group was observed in males, while a significant decrease in concentration of the low ω-6/ω-3 ratio found in the FO group was observed in females. Low dietary Zn intake was observed in both male and female rabbits fed DHA/ARA or FO diets as a consequence of low food intake. During Zn deficiency, the amount of this mineral in bone decreases because bone is an endogenous source of Zn when the dietary supply is low [31]. The mechanism underlying the different response in femur Zn content to low ω-6/ω-3 ratio between males and females found in the current study is unclear and further investigation is needed; however, it suggests that Zn metabolism is altered in response to LCPUFA, particularly to their ratio.
In the current study, results of regression analysis showed that as the dietary ω-6/ω-3 ratio was elevated, the level of PGE2 rose, while Ca, P, and Mg contents in femur decreased. In addition, as the dietary ARA/EPA ratio increased, the PGE2 level and that of all of these minerals increased. In the current study, the favorable effect of ARA/EPA ratio on Ca, P, and Mg contents suggests that the particular bioactive fatty acid concentration and their ratio per se in our oils blend may be more important to bone growth than the overall ω-6/ω-3 ratio in the diet. Of particular interest, femur Zn content increased with elevation of both ratios. In addition, the results of the current study showed that as the level of PGE2 was elevated, Ca, P, and Mg contents in the femur decreased while Zn content increased. The mechanism by which the elevation of PGE2 decreased Ca, P, and Mg and increased Zn content in femur in the current study is unclear; however, measurements of mineral absorption and balance were not performed in this study and could potentially explain this conflicting effect of PGE2 on bone mineral content.
In conclusion, the current study clearly demonstrated that dietary PUFA, particularly ω-6/ω-3 and ARA/EPA ratios are important factors in determining bone marrow fatty acid profile, and this in turn determines the capacity of bone for synthesis of PGE2, thereby reducing bone resorption and improving bone mass during growth. The reduction in the ω-6/ω-3 ratio resulted in a significant increase in femur Ca, P, and Mg contents in both genders, although low dietary intake of these minerals suggests an important role of LCPUFA in mineral metabolism and bone mineralization. In addition, the reduction in the ω-6/ω-3 ratio using fish oil or marine algae oils as sources of ARA, EPA, and DHA supports femur Ca, P, Mg, and Zn contents depending on the dietary amount of these fatty acids and, more importantly, their ratio. In addition, the significant elevation in mineral content and the maintenance of optimal Ca/P ratio in bones of DHA/ARA and DHA fed groups has proven that marine algae oils may be promising dietary sources for promotion of bone mineralization during the growing stage.





 XML Download
XML Download