

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Hereditary breast cancers occur in about 5% to 10% of patients with breast cancers [12]. Germline mutations occurring in the BRCA1 and BRCA2 genes have been identified as the causal mutations for hereditary breast, ovarian, and other related cancers [34]. Carriers of BRCA1 mutations are estimated to have lifetime risks of 87% and 44% for breast and ovarian cancers, respectively [56]. The most pathogenic mutations in the BRCA1 and BRCA2 genes are the nonsense mutations and the small insertions and deletions, which lead to the translation of truncated proteins [78]. Furthermore, splicing analysis of the BRCA1 and BRCA2 variants has revealed that several mutations at the exon-intron boundaries disrupt the accuracy of splicing of the corresponding mRNAs, resulting in altered splicing events [910]. Point mutations and nonsense mutations in the BRCA1 gene that cause the skipping of one or more exons have been reported in patients with cancers [11]. Here, we report a novel germline mutation in the BRCA1 gene that results in exon 20 skipping in a family with a history of breast and ovarian cancers.
CASE REPORT
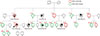
A 43-year-old woman with breast cancer visited the National Cancer Center in Korea for obtaining treatment, and she was referred to the genetic counseling clinic, due to her family history of breast and ovarian cancers. Her pedigree was assessed, and blood samples were collected from 17 family members. The proband had been diagnosed with breast cancer at 43 years of age, and her mother had died of ovarian cancer at an age of 66 years. Among the maternal relatives of the proband, two cousins had been diagnosed with breast cancers, but further information was not available (Figure 1). The proband underwent nipple-sparing mastectomies of both breasts, and immunohistochemistry revealed that her tumor tissue was of a triple negative breast cancer subtype, i.e., negative for estrogen and progesterone receptors and for human epidermal growth factor receptor 2. Despite strong recommendations by her doctors, she refused an oophorectomy, but she did undergo regular gynecological examinations, including trans-vaginal ultrasounds.
The genetic testing of the BRCA1 and BRCA2 genes in the proband was performed by using a direct sequencing method. Genetic testing of the proband showed a single nucleotide deletion in the BRCA1 intron 20, namely c.5332+4delA (Figure 2A). To test the mRNA sequence of BRCA1, total RNA was extracted from proband's blood, and the cDNAs corresponding to BRCA1 exons 19, 20, and 21 were sequenced. RNA sequencing showed that the abovementioned deletion had resulted in exon 20 skipping and the introduction of a premature stop codon (Figure 2B). The mother of the proband had died of ovarian cancer and had harbored the same mutation. Among 17 other family members, two uncles and two cousins too harbored the same mutation.
We examined whether the mutation had a functional effect on the BRCA1 protein. Mutant (exon 20 deletion) and wild-type (WT) BRCA1 were overexpressed in a breast cancer cell line, MDA-MB-436 cells, and the resultant protein expression levels were estimated by Western blotting. Exogenous expression of these proteins in MDA-MB-436 cells revealed that the levels of the mutant protein were much lower than those of WT BRCA1. Following treatment of the transfected cells with MG132 to block potential proteasomal degradation, the levels of mutant and WT proteins in these cells were comparable (Figure 3A). To assess whether mutant BRCA1 could interact with BARD1, BRCA1, and BARD1 proteins were co-immunoprecipitated as described previously [12]. Both the WT and mutant proteins immunoprecipitated with BARD1 (Figure 3B), in proportion to their level of expression. The levels of cyclin B1 mRNA in these cells were measured by real-time reverse transcription polymerase chain reaction, following the exogenous expression of WT or mutant BRCA1. WT BRCA1 enhanced the expression levels of cyclin B1 mRNA; however, in cells that overexpressed mutant BRCA1 with or without WT, the levels of cyclin B1 mRNA remained unchanged (Figure 3C).
DISCUSSION
Germline mutations of the BRCA1 and BRCA2 genes, which result in truncated proteins, have been identified as the pathogenic mutations that cause the hereditary cancers. Carriers of the BRCA1 and BRCA2 mutations should be monitored carefully, due to the greatly increased risks of the incidence of cancers in these individuals.
Here, we report a patient who harbored a single nucleotide deletion in intron 20 of the BRCA1 gene. The mother of this patient had ovarian cancer and harbored the same mutation. Among 17 other family members, two uncles and two cousins too harbored the mutation.
A single nucleotide deletion at the 5' end of intron 20 caused an altered splicing event, which resulted in the production of an altered sequence beginning at the amino acid at position 1760 in the sequence; this, in turn, resulted in the translation of a truncated protein with 1773 amino acids. A truncated BRCA1 protein with a short C-terminal sequence may display an abnormal protein lacking the BRCA1 C-terminal (BRCT) domain, which has been reported to be essential for the tumor suppressor function and protein interaction of BRCA1 [13]. Splicing variants in exons 19–21 of the BRCA1 gene that cause exon skipping or intron retention due to aberrant splicing have been reported in patients with breast cancers [1014].
Functional analyses of the mutant BRCA1 protein (exon 20 deletion) revealed that the protein levels of this mutant were lesser than those of the WT BRCA1. Furthermore, the results of inhibiting proteasomal degradation following the exogenous expression of these proteins suggested that the mutant protein was more susceptible to proteasomal degradation. Results of immunoprecipitation studies showed that both the mutant and WT BRCA1 could interact with BARD1. However, the mutation impaired the transcriptional activity of BRCA1. To assess the effect of the c.5332+4delA mutation on the transcriptional function of BRCA1, the levels of cyclin B1 mRNA were compared between cells that exogenously expressed the WT and mutant BRCA1 proteins. Consistent with previous reports [15], the WT BRCA1 altered the expression of cyclin B1 mRNA. However, the mutant BRCA1 could not alter the levels of cyclin B1 mRNA (Figure 3C). Interestingly, cells expressing both WT and mutant BRCA1 showed normal levels of cyclin B1 mRNA, suggesting that the mutant protein has a dominant negative effect.
In conclusion, this report describes a novel germline mutation of BRCA1 that causes exon 20 skipping in a Korean family with a history of breast cancer. These data suggest that germline mutations occurring in splice site consensus sequences should be investigated carefully in families with histories of cancers.


 XML Download
XML Download