

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Melanoma develops from melanocytes, which produce the pigment melanin in the skin and oral mucosa. Melanoma occurs when melanocytes change and grow out of control. It is the most aggressive and malignant form of skin cancer.1 The treatment of melanoma involves a combination of surgery, radiation treatment, and chemotherapy.2 However, surgical treatment involves painful tissue resection or skin grafting, and the success of chemotherapy is hindered by the frequent development of resistance. A conservative, non-invasive technique for the selective induction of apoptosis in cancer cells is needed.
Nanoparticles (NPs) have become recognized as an intracellular transport system for many components, such as nucleic acids and proteins. Nanotechnology has been applied to various fields, including for the prevention, diagnosis, and treatment of cancer.3 The use of NPs for intracellular transportation exploits benefits of NPs that include size, form, biosafety, intracellular absorption, and intracellular dispersion. Effective cancer cell responses using NPs have included the use of carbon-nanotubes,456 calcium-selenide-NPs,78 ZnO-nanorods,9 chitosan-NPs,1011 and gold NPs.1213
Research on the biosafety of gold in humans spans half a century. As a targeted drug delivery agent, gold has never been reported to show cytotoxicity.1214 In the medical field, gold NPs (GNP) are unevenly dispersed colloidal particles, with a wide surface area that ranges from a few nanometers to hundreds of nanometers. GNPs are biocompatible and non-toxic, are easily synthesized, and combine well with biological materials. These benefits have made GNPs a recognized alternative for treatments, including cancer. GNPs have been reported to combine with some cancer cells to induce apoptosis without special surface treatment.15 GNPs easily combine with many biological ligands, such as a single antibody or a peptide, which can be used as a probe to selectively recognize cancer.1617 The combination of GNP and antibody is stable and can effectively enter cells. In particular, GNP combined with antibody to epithelial growth factor receptor (EGFR), which can be present on the surface of cancer cells, is very useful for imaging cancer by causing an antigen-antibody reaction in squamous epithelium cell carcinoma.131516
HER2 is a cancer marker and is one of the four components in the EGFR superfamily: HER1 (ErbB1 or EGFR), HER2 (ErbB2), HER3 (ErbB3), and HER4 (ErbB4). HER2 is a 185 kDa cell membrane glycoprotein composed of intracellular tyrosine kinase region and extracellular ligand binding region.1718 HER2 controls the growth, survival, adhesion, and migration of cells. It accompanies cell signal transduction pathways that control cancer cell amplification or weakening.1920 The over-expression of HER-2 can affect cancer malignancy in many regions of the body, such as in breast, ovary, lung, uterus, and stomach.21222324 GNP combined with the antibody against HER2 (GNP-HER2) is non-toxic to normal cells, and very avidly and selectively conjugates with melanoma cells.
Melanoma cells combined with GNP-HER2 polymer display functional abnormalities and induction of cell death. Natural cell death during apoptosis is one form of cell death by gene control, and is distinguished from cell necrosis.2526 When natural cell death occurs, the cell dies from cell contraction by dehydration, DNA fragmentation, chromatin condensation, activation of caspase, cell cycle arrest, and formation of an apoptotic body.27
This study was conducted to verify how synthesized GNP-HER2 polymer specifically targets melanoma cells and induces death and to investigate the mechanism of selective apoptosis.
MATERIALS AND METHODS
Cell culture
G361 and HaCaT cells were cultured in Dulbecco's modified Eagle's medium (Gibco, New York, NY, USA) supplemented with 10% heat-inactivated fetal bovine serum (FBS, Gibco), 4 mM L-glutamine, and 100 µg/mL of penicillin/streptomycin at 37℃ in a humidified incubator with a 5% CO2 atmosphere.
GNPs conjugated with antibodies
An aqueous solution of 11-mercaptoundecanoic acid (MUA) (0.1 mg/mL) was added to the colloidal gold suspension and incubated overnight. MUA-modified GNPs reacted with a mixture of 1 mM N-hydroxysuccinimide (NHS) and 1 mM N-ethyl-N'-(3-dimethylaminopropyl) carbodiimide (EDC) solution for 20 min. NHS-terminated GNPs were incubated with HER 2 antibody in phosphate buffered saline (PBS; 1 mM, pH 7.0) for >8 h (Fig. 1A).
Cell viability assay
In vitro cell viability was evaluated by the WST-1 assay. Cells (1×104) were seeded in wells of a 96-well plate. The cells were treated with GNP-HER2. After 24, 48, and 72 h incubation, 10 µL of WST-1 reagent was added to each well, and absorbance at 450 nm was measured after 2 h using a micro plate reader (Sunrise Remote Control, Tecan, Austria). The assay was performed in triplicate.
Hemacolor staining
Cells were spread on a cover glass, air-dried, and immersed in 4% paraformaldehyde. These slides were immersed in red and blue reagent solution. The cells were washed twice with PBS and mounted in 100% glycerol. The morphological characteristics of the cells were determined using optical microscopy (Zeizz Axioskop, Jena, Germany).
Nuclear staining with Hoechst 33258
Cells were incubated for 24 h and harvested by cyto-centrifugation. They were then washed twice in PBS and incubated with 5 µg/mL of Hoechst 33258. The morphological characteristics of apoptotic cells were observed with the aid of an LSM 700 laser-scanning confocal microscope (Carl Zeiss, Göettingen, Germany).
Immunocytochemistry
Cells were treated with GNP-HER2 and fixed in 4% PFA for 5 min. Cells were permeabilized with 0.1% Triton X-100 in PBS for 10 min at 4℃ and then incubated with goat Alexa 488 anti-mouse secondary antibody for 60 min. Fluorescent images were observed and analyzed using the aforementioned laser-scanning confocal microscope.
Western blot analysis
Cells were lysed with lysis buffer (10 mM Tris/HCl, pH 7.2, 1% Triton X-100, 150 nM NaCl, 5 mM EDTA) on ice for 1 h. The lysates were clarified by centrifugation at 14000 rpm for 20 min at 4℃, and the supernatant was obtained. The protein content of the lysate was determined using a protein assay kit (Bio-Rad Laboratories, Hercules, CA, USA). The samples (50 µg of lysate) were boiled for 95℃ for 5 min, and the protein was resolved by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to polyvinylidene fluoride membranes. After transfer, the membranes were blocked with a blocking reagent [5% non-fat milk in TBS-T (20 mM Tris, 150 mM NaCl, 0.1% Tween 20)] for 1 h. The membranes were incubated for 2 h with the corresponding antibody. The membranes were treated with ECL Western blotting reagents and detected.
Flow cytometry analysis
The G361 cells were seeded in wells of a 6-well plate at a density of 1×105 cells/well and incubated for 24 h. GNP-HER2 treated cells were incubated for the same time. Cells were harvested, washed with cold PBS, centrifuged at 1500 rpm for 5 min, and added to cold 70% ethanol for 24 h. The fixed cells were washed with PBS and centrifuged at 1500 rpm for 5 min. They were incubated in the presence of RNAase (100 µg/mL) at 37℃ for 30 min and resuspended in propidium iodide (PI) solution (10 µg/mL). Cells were incubated at 4℃ for 10 min and analyzed using a FACS Canto II apparatus (BD Biosciences, San Jose, CA, USA).
RESULTS
Comparison of HER2 protein expression between melanoma cells and a normal cornified cells
To determine if HER2 protein is a selective marker for G361 cells, HER2 expression was compared between G361 melanoma cells and HaCaT normal cornified cells using the Western blot analytical method. HER2 was expressed at a markedly higher level in G361 cells than in HaCaT cells (Fig. 1B).
Selective induction of cancer cell apoptosis by GNP-HER2 polymer
To observe the influence of GNP-HER2 on the survival of G361 and HaCaT cells, WST-1 analysis was performed. G361 cell viability was reduced to 69% at 24 hours of GNP-HER2 treatment, 8% at 48 hours, and 2% at 72 hours (Fig. 2A). On the other hand, HaCaT cell viability increased to 126% at 24 hours, 124% at 48 hours, and 86% at 72 hours (Fig. 2B). Also, when only the antibody against HER2 was used for treatment, cell viability decreased (110% at 24 hours, 54% at 48 hours, and 11% at 72 hours). Compared to treatment with GNP-HER2 polymer, apoptosis was markedly reduced (Fig. 2C). GNP-HER2 reduced the viability of melanoma cancer cells more rapidly than when an antibody against HER2 was used alone. The viability of normal cells was not affected.
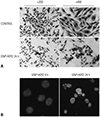
Morphological change of G361 cells following GNP-HER2 treatment
To observe morphological changes in G361 cells upon GNP-HER2 treatment, Hemacolor and Hoechst staining was performed. The control G361 cells displayed round nuclei and a normal, spindle-shaped cell membrane. However, after 24 hours of GNP-HER2 treatment, nuclei were condensed, and rupture of cell membranes was evident (Fig. 3A). The G361 control cells displayed round nuclei; however, compared to the cells treated for 24 hours with GNP-HER2, nuclei were condensed and disassembled apoptotic bodies were observed (Fig. 3B). The morphological changes of cancer cells were indicative of cell apoptosis.
Positional changes of apoptosis inducing factor and cytochrome c
Immunofluorescence staining was examined using confocal microscopy to observe the positional changes of apoptosis inducing factor (AIF) and cytochrome c. These changes are related to mitochondria undergoing apoptosis. In the control group, AIF and cytochrome were located within the mitochondria. However, in the group treated with GNP-HER2, AIF moved from the mitochondria to the nucleus, and cytochrome c moved from the mitochondria to the cytoplasm (Fig. 4).
Activation of proteins related to apoptosis
The effect of the B-cell lymphoma 2 (Bcl-2) family associated with typical apoptosis signal transduction was observed using Western blot analysis. In cells treated with GNP-HER2, BAX increased with time and Bcl-2 decreased (Fig. 5A). Caspase-3 and -9, which are typical apoptosis-inducing control elements, displayed 17 kDa and 34 kDa cleavage forms at 12 hours and were activated with time (Fig. 5B). Due to the activation of caspase-3, poly (ADP-ribose) polymerase (PARP), which is involved in DNA repair, and DNA fragmentation factor 45 [inhibitor of caspase-activated DNase (ICAD, DFF45)], which inhibits DNase DFF40, were progressively destroyed with time, indicating the acceleration of cell apoptosis (Fig. 5C).
Sub-G1 index determination by fluorescence-activated cell sorting analysis
Flow cytometry and Western blot analysis were performed to evaluate changes in sub-G1, which is related to each cell apoptosis, and the amount of protein. In flow cytometry, PI was used as a DNA stain, since it binds to DNA and emits fluorescence: it measures sub-G1 DNA content within the cell and shows the percentage of apoptotic cells. Following GNP-HER2 treatment, an increase in sub-G1, an indicator of cell apoptosis, was observed, compared to the control group (0 h: 1.6%, 12 h: 6.9%, 24 h: 28.6%, 48 h: 49.2%) (Fig. 6).
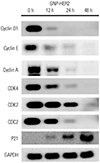
Changes in protein amount in relation to the cell cycle
Western blot analysis was performed to investigate changes in protein amount in relation to the cell cycle. After GNP-HER2 treatment, cyclin D1 and cyclin-dependent kinase 4 (cdk4), which are related to the G1 phase, were down-regulated. Cyclin A, cyclin E, cdk2, and cdc2, which are related to S and G2/M phase, were also down-regulated with time. In addition, p21, which is known to induce the synthesis of cdk inhibitory protein and inhibit cdk activation, continuously increased beginning at 12 hours (Fig. 7).
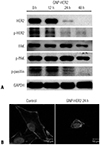
Changes in protein amount of focal adhesion kinase and F-actin disassembly
Western blot analysis and immunofluorescence staining were performed to investigate changes in protein amount of focal adhesion kinase (FAK) and F-actin disassembly. After GNP-HER2 treatment, the amount of FAK, which plays a significant role in focal adhesion formation, decreased with time. FAK phosphorylation levels, which indicate an activated state, were also decreased. Furthermore, the amount of HER2 protein, which is a target of GNP-HER2, and HER-2 phosphorylation (p-HER2) decreased. Phosphorylation of paxillin, which is a downstream signal transduction protein of HER2 with an important role in actin synthesis, decreased as well (Fig. 8A). Compared to the control group, following the 24-hour GNP-HER2 treatment, focal adhesion was disassembled, as was the filamentous actin cellular structure, which led to an extreme cell contraction (Fig. 8B).
DISCUSSION
Recently, cancer treatment utilizing a combination of immune cells and NPs or the use of exothermic technology with GNPs during photothermic treatment of cancer cells has become popular. However, the anticancer mechanisms of NPs are unclear.
In this study, the conjugation between the most expressed protein in G361 melanoma cells treated with GNPs and subsequent apoptosis was investigated and compared with normal cells. Comparison of HER2 expression between G361 and HaCaT cells confirmed the marked increase of HER2 in G361 cells (Fig. 2A). GNP-HER2 treatment markedly decreased cell proliferation and survival rates with time (Fig. 2A). In normal HaCaT cells, proliferation and survival were unaffected. When HER2 antibody was omitted, apoptosis at 24 hours was markedly less, even though survival was observed at 48 hours. GNP-HER2 polymer attached well to HER2, and the anticancer effects of GNPs and of HER2 antibody were synergistic. Therefore, it is assumed that the selectively induced death of melanoma cells was more dominant.
Natural death in cells plays a significant role in managing individual development, as well as functional sustainability. It is considered an important target in the treatment of cancer. The Bcl-2 family is very important in regulating the intrinsic pathway. When the balance is disturbed between them, AIF28 is released and enters the nucleus. Cytochrome C29 flows into the cell transforming into Apaf1/cytc/ATP, thus activating caspase-9 and causing cell natural death through mechanical failure of the mitochondria. Both routes cause cell death by activating caspase-3. We analyzed how various proteins participating in the intrinsic apoptotic pathway influence natural cellular death upon treatment with GNP-HER2. AIF, which is normally localized in mitochondria was translocated into the cytoplasm and then the nucleus in GNP-HER2 treated cells. This phenomenon reveals that natural cellular death occurs upon GNP-HER2 treatment. Furthermore, it was verified that cytochrome c, which resides in mitochondria was translocated into the cytoplasm in the GNP-HER treated group. The Bcl-2 family, including Bcl-2, Bax, Bak, etc., is the protein group that controls the stimulation and suppression of natural cellular death and is localized in the mitochondrial inner membrane, cytoplasm, and endoplasmic reticulum.30 Bcl-2 and Bcl-XL block cytochrome c release, hindering natural cellular death, and Bid, Bax, and Bak hasten cytochrome c release, which induces natural cellular death. When the most representative apoptosis suppression signal Bcl-2 and stimulant signal BAX were analyzed by Western blot analysis in this experiment, we decreases in Bcl-2 and increases in BAX in a time-dependent manner.
Caspase is divided into initiator caspases (caspase-8 and -9) and effector caspases (caspase-3, -6, and -7). When caspases are activated, the downstream caspase cleaves several types of substrate proteins, inducing natural cellular death. We examined the effect of GNP-HER2 on the expression and activity of caspase-3 and -9, which are directly involved in apoptosis. The level of caspase-3 was increased as was the level of caspase-9 by GNP-HER2 treatment. In addition, expressions of PARP31 and DFF45 (ICAD),32 which are substrate proteins of caspase-3, were reduced or fragmented by GNP-HER2 treatment. These results suggest that caspase-3 plays a more important role in the final stage of apoptosis, although several proteins are involved in cellular apoptosis induced by GNP-HER2. The structures formed by integrin binding to extracellular matrix are called focal adhesions, and these structures serve as a starting point for signaling mediated by integrin. In addition, focal adhesion plays a crucial role in cell shape changes related to cell migration, attachment, and spreading. These focal adhesion sites comprise various proteins, such as FAK, paxillin, talin, vinculin, tensin, and p130cas. When focal adhesions are destroyed, cells lose contact with the extracellular matrix and die. On analyzing the proteins involved in focal adhesions during G361 cell death, a gradual decrease in the amounts of FAK, HER2, paxillin, and phosphorylated proteins was observed. As a result of treatment with GNP-HER2, primary changes in cell shape during cell death were observed, and cell shrinkage, as well as the collapse of filamentous actin and destruction of cell adhesion, were observed.
These results suggest that treatment of G361 and HaCaT cells with GNP-HER2 does not affect normal keratinized HaCaT cells, whereas a reduction in cell viability and morphological changes were observed in melanoma G361 cells and were closely related to cell death. Activation of caspase-9 and -3 by degradation of Bcl-2 and substrate proteins, inhibition of cell cycle, and focal adhesion and cytoskeleton destruction were involved in induction of apoptosis caused by GNP-HER2 treatment. Therefore, it can be concluded that GNP-HER2 treatment selectively induces apoptosis in melanoma cells without affecting normal cells. Because mutated HER2 genes are observed in some melanoma cells, we wonder if HER2-GNP could also induce the selective killing effect on such melanoma cells. Thus, further study is needed to investigate the affinity between HER2-GNP and mutated HER2.





 XML Download
XML Download