

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Mast cells are bone marrow-derived effector cells of the immune system, found abundantly in connective tissue, skin, mucosal membranes, and tissues which interface with the external environment. It has been thought that mast cells play a major role in the development of many physiological changes during allergic responses. Mast cell activation by both IgE-dependent and IgE-independent stimuli initiate degranulation which results in the fusion of cytoplasmic granule membranes with plasma membranes. This process is accompanied by the fast external release of granule-associated stored mediators (histamine, neutral proteases, acid hydrolyases, proteoglycans, chemotactic factors, cytokines such as tumor necrosis factor-α, interleukins, etc.) as well as the generation and release of newly generated mediators, including products of arachidonic acid metabolism and an array of cytokines (Galli & Tsai, 2010). Histamine, a major component of mast cell granules, exerts many effects related to the immediate-phase of allergic inflammation including vasodilation, increased vascular permeability, tissue edema, contraction of bronchial and intestinal smooth muscle, and increased mucus production (Galli et al., 2008). Compound 48/80 is known to be a potent inducer of degranulation, responsible for the release of histamine and other chemical mediators associated with anaphylactic symptoms, and the activation of mast cells. Compound 48/80 also causes a rapid release of inflammatory substances only from connective tissue-type mast cells, specifically rat peritoneal mast cells (RPMCs) (Nishikawa & Kitani, 2008). Thus, RPMCs are a good experimental model for the study of compound 48/80-induced histamine release and anaphylaxis-like reactions.
Alpha-lipoic acid (1,2-dithiolane-3-pentanoic acid; LA), a disulphide derivative of octanoic acid, is known to act as an efficient anti-oxidant and metal-chelating agent (Shay et al., 2009). LA has been used to treat many diseases including diabetic complications, polyneuropathies, atherosclerosis, diseases of joints, and acquired immune deficiency syndrome (Bilska & Wlodek, 2005). LA has also been considered as a therapeutic agent candidate for the treatment or prevention of pathologies associated with an imbalance of oxidoreductive status, such as neurodegeneration, ischemia-reperfusion, hepatic disorders, and asthma (Shay et al., 2009). Recent studies have revealed that LA effectively suppresses allergic inflammation in a murine model of asthma by reducing the level of reactive oxygen species (Cho et al., 2004). However, the effects of LA on mast cell-mediated allergic inflammation and cellular regulation remain unknown.
In this study, we aimed to investigate the effect of LA on mast cell-mediated immediate-type hypersensitivity. The results clearly demonstrated that LA displays anti-allergic activity in both in vivo and in vitro models.
Materials and Methods
Materials
Lipoic acid (LA; Thioctacid 600) was obtained from VIATRIS GmbH & CoKG (Frankfurt, Germany). Compound 48/80, bovine serum albumin (BSA), disodium cromoglycate (DSCG) and HEPES were purchased from Sigma Chemical Co. (St. Louis, MO, USA). Percoll solution was purchased from Pharmacia (Uppsala, Sweden). LA and compound 48/80 were freshly diluted in phosphate buffered saline (PBS) for all in vitro experiments.
Experimental animals
Male ICR mice and Sprague Dawley rats were purchased from Damool Science (Daejeon, Korea). Animals were housed 3~5 per cage in laminar air-flow cabinets maintained at 22±1℃ and a relative humidity of 55±10% throughout the study. All experiments were performed in compliance with the guidelines approved by the Institutional Animal Care and Use Committee of the Chonbuk National University Medical School.
Compound 48/80-induced systemic anaphylaxis-like reactions in mice
Mice (n=10/group) received an intraperitoneal injection of compound 48/80 (8 mg/kg body weight [BW]) as described previously (Choi et al., 2006). LA (10, 50, 100 mg/kg BW) was diluted in PBS and administered orally at 24, 12, and 1 h prior to compound 48/80 administration. Mortality was monitored for 1 h after induction of anaphylactic shock. At the conclusion of the mortality test, blood was obtained by cardiac puncture.
Preparation of plasma and histamine determination
The blood was centrifuged at 150×g for 10 min at 4℃ and histamine content was measured by radioenzymatic method from the plasma (Carvalho et al., 2010).
Compound 48/80-induced a passive cutaneous anaphylaxis (PCA)-like reaction
After intradermal injection of 0.5 µg/20 µl of compound 48/80 to the dorsal skin of each mouse, 2% Evans blue solution was intravenously injected into the lateral tail vein. LA was administered orally 24, 12, and 1 h before the injection of compound 48/80. As a control, a mast cell-stabilizing agent, DSCG, was given orally 24, 12, and 1 h prior to the injection of compound 48/80. Mice were euthanized with a terminal anesthesia dose 30 min after the 2% Evans blue injection. Tissue sections around the intradermal injection site were excised and weighed. Extravasated Evans blue dye was extracted by incubation of biopsies in 1 ml formamide at 55℃ for 24 h. Absorbance was measured at 620 nm using a spectrophotometer (Spectra MAX PLUS, Molecular Devices, CA, USA). Tissue Evans blue concentrations were quantified by interpolation on a standard curve of dye concentrations in the range of 0.01 to 30 µg/ml.
Preparation of mast cell suspensions and microscopic observation
RPMCs were isolated as follows: rats were anesthetized with ether and injected with 10 ml of calcium-free HEPES-Tyrode's buffer (136 mM NaCl, 5 mM KCl, 11 mM NaHCO3, 0.6 mM NaH2PO4, 5.4 mM HEPES, 1.0 mg/ml glucose) into the peritoneal cavity, and the abdomen gently massaged for about 90 s after which the peritoneal cavity was opened and the fluid aspirated using a Pasteur pipette. RPMCs were purified using a Percoll density gradient as described in detail elsewhere (Martynova et al., 2005). RPMC preparations were at least 95% pure as assessed by toluidine blue staining and at least 98% of these cells were viable according to trypan blue exclusion. Purified mast cells (1×106 cells/ml) were resuspended in HEPES-Tyrode's buffer (136 mM NaCl, 5 mM KCl, 2 mM CaCl2, 11 mM NaHCO3, 0.6 mM NaH2PO4, 2.75 mM MgCl2, 5.4 mM HEPES, 1.0 mg/ml BSA, 1.0 mg/ml glucose, 0.1 mg/ml heparin, pH 7.4), observed under a phase contrast microscope and photographed.
Mast cell viability assay
To test the viability of cells, the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) colorimetric assay was performed as described previously (Yoshimura et al., 2004). Briefly, RPMCs (1×106 cells/well) were incubated with various concentrations (1~4 mM) of LA at 37℃ for 2 h. After addition of MTT (100 µg in 100 µl saline) cells were incubated at 37℃ for 1 h. The crystallized MTT was dissolved and absorbance measured at 570 nm with a spectrophotometer.
Histamine release assay
Mast cell suspensions (2×105 cells in 200 µl) were preincubated with LA (1~4 mM) at 37℃ for 10 min and then incubated with compound 48/80 (0.25 µg/ml) for 10 min. Following centrifugation at 150×g for 10 min, the amount of histamine in the supernatant was determined by the radioenzymatic method (Carvalho et al., 2010). The inhibition percentage of histamine release was calculated using the following formula:
Measurement of 45Ca uptake
Purified mast cells were resuspended in HEPES-Tyrode's buffer containing 45Ca (1.5 mCi/ml; 1 Ci=3.7×1010 becquerels; Perkin-Elmer Life Sciences, MA, USA) and incubated at 4℃ for 10 min. Mast cell suspensions were preincubated with LA (1~4 mM) at 37℃ for 10 min and then incubated with compound 48/80 (0.25 µg/ml) for 10 min. The reaction was stopped by the addition of 1 mM lanthanum chloride and samples were centrifuged 3 times at 150×g for 10 min at 4℃. Mast cells were then lysed with 10% Triton X-100 and vigorous shaking. Radioactivity of the solution was measured in a scintillation β-counter (Liquid Scintillation Analyzer, A Canberra Company, Australia).
Measurement of cyclic adenosine-3',5' monophosphate (cAMP) levels
Mast cell suspensions were added to an equivalent volume (200 µl) of prewarmed buffer containing LA (1~4 mM) in Eppendorf tubes. The reaction was allowed to proceed for specific time intervals, terminated by centrifugation. Each sample was then added to 250 µl of 50 mM sodium acetate buffer (pH 6.2) under vigorous vortexing, followed by snap freezing in liquid nitrogen. Frozen samples were thawed and vortexed, and the debris sedimented by centrifugation 1,200×g at 4℃ for 10 min. The cAMP levels in the supernatants were determined with cAMP radioimmunoassay kits (Perkin-Elmer Life Sciences).
Results
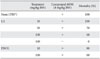
Effect of LA on compound 48/80-induced systemic anaphylaxis-like reactions
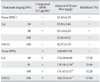
To investigate the effect of LA on anaphylaxis-like reactions, we first used a mouse in vivo model with compound 48/80. After intraperitoneal injection of compound 48/80 (8 mg/kg BW) the mortality rate was examined for 1 h. As shown in Table 1, injection of compound 48/80 resulted in 100% death. Oral administration of LA (10, 50, 100 mg/kg BW) reduced compound 48/80-induced mortality in a dose-dependent manner. DSCG (reference drug) also dose-dependently inhibited compound 48/80-induced mortality.
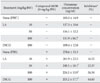
Effect of LA on compound 48/80-induced plasma histamine release
LA was given at doses ranging from 10 to 100 mg/kg BW at 24, 12, and 1 h prior to injection of compound 48/80. Oral administration of LA reduced the plasma histamine release induced by compound 48/80 in a dose-dependent manner (Table 2). In addition, the reference drug DSCG attenuated compound 48/80-induced plasma histamine release.
Effect of LA on compound 48/80-induced PCA-like reactions
It has been previously demonstrated that intradermal injection of compound 48/80 into the dorsal skin of mice provokes the increase of mast cell-dependent vascular permeability (Shinmei et al., 2004). As shown in Table 3, oral administration of LA dose-dependently reduced the vascular permeability changes induced by compound 48/80. Likewise, DSCG significantly inhibited the PCA-like reaction triggered by compound 48/80 at a dose of 100 mg/kg BW.
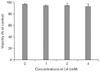
Effect of LA on mast cell viability
An MTT conversion assay was used to determine the viability of RPMCs exposed to LA. Cell viability was almost 100% after exposure to 4 mM LA for 2 h (Fig. 1); thus, LA had no cytotoxic effects on RPMCs.
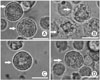
Effect of LA on mast cell degranulation of RPMCs
To investigate the inhibitory mechanism of LA on anaphylactic reactions, we examined compound 48/80-induced mast cell activation (Fig. 2). Control RPMCs were generally spherical or oval in shape and contained many fine granules surrounding a prominent nucleus (Fig. 2A). Following stimulation with compound 48/80 for 10 min, RPMCs showed characteristics of mast cell degranulation with cell swelling, cytoplasmic vacuoles, and extruded granules near the cell surface and in the surrounding medium (Fig. 2B). When RPMCs were incubated with LA alone they appeared similar to control cells (Fig. 2C). Pretreatment with LA inhibited degranulation of RPMCs induced by compound 48/80. The cell sizes were somewhat larger than the control cells (Fig. 2D); however, there was no statistical significant difference in size between the two groups (data not shown). These results suggest that LA suppresses compound 48/80-induced mast cell degranulation.
Effect of LA on histamine release from RPMCs
Histamine release from compound 48/80-treated RPMCs was reduced in a dose-dependent manner by LA (Fig. 3). DSCG also suppressed compound 48/80-induced histamine release. These results indicate that LA inhibits the compound 48/80-induced anaphylactic-like reaction by blocking histamine release from RPMCs.
Effect of LA on calcium uptake into RPMCs
Calcium plays an important role in the stimulus-secretion coupling of mast cells. It is also well established that an increase in calcium uptake into RPMCs contributes to the release of histamine (Akagi et al., 1994). Therefore, we measured calcium uptake and found that treatment with LA had no effect on calcium uptake. However, calcium uptake was substantially increased by stimulation of RPMCs with compound 48/80. This compound 48/80-elicited calcium uptake response was inhibited by LA in a concentration-dependent manner (Fig. 4). At a concentration of 4 mM of LA, calcium uptake was nearly completely blocked. These results suggest that LA may inhibit mediator release by blocking calcium uptake into RPMCs.
Effect of LA on cAMP levels of RPMCs
Earlier findings have shown that the cAMP pathway participates in the modulation of mast cell activation. Increased cAMP is known to precede the prevention of mediator release from mast cells activated by compound 48/80 or IgE (Tasaka et al., 1986). To investigate the mechanism of LA involved in the reduction of mediator release from RPMCs stimulated by compound 48/80, we assessed intracellular cAMP levels. The level of cAMP was not changed in unstimulated RPMCs. Treatment of RPMCs with LA resulted in an increase in cAMP in a concentration-dependent manner. When RPMCs were incubated with 2 or 4 mM LA, the cAMP level was rapidly increased within 1 min and decreased to the basal level after 3 min (Fig. 5). These results suggest that LA may suppress mediator release by increasing the cAMP level in RPMCs.
Discussion
The present study has demonstrated that LA inhibits compound 48/80-induced systemic anaphylaxis-like and passive cutaneous anaphylaxis-like reactions in mice. Pretreatment with the same concentration of LA before the systemic anaphylaxis-like reaction also reduces the plasma histamine content in a dose-dependent manner. To further examine the mechanism by which LA attenuates mast cell-mediated anaphylactoid reactions, we examined compound 48/80-induced mast cell activation. Compound 48/80-induced histamine release from RPMCs was blocked by treatment with LA. Compound 48/80 is well-known to induce mast cell-dependent, non-specific anaphylactic reactions. Additionally, it is widely accepted that the mechanism involved in the anaphylaxis-like response activated by compound 48/80 is due to the massive release of histamine from mast cells and basophils (Nishikawa & Kitani, 2008). Therefore, these results suggest that LA could inhibit the mast cell-derived anaphylactoid reactions by preventing histamine release from RPMCs triggered by compound 48/80.
Several reports have shown that compound 48/80 increases the permeability of the lipid bilayer membrane of mast cells by causing perturbation of the membrane (Tasaka et al., 1986). These findings indicate that the increase in cell membrane permeability may be an essential trigger for the release of mediators from mast cells. Thus, it is reasonable to presume that LA might inhibit histamine release by attenuating the permeability of RPMC membranes by preventing compound 48/80-induced membrane perturbation. However, we found no evidence in our study to substantiate this assumption. In order to reveal the mechanisms involved in histamine release inhibition by LA, additional research is required.
To further examine the mechanisms by which LA inhibits mast cell-dependent anaphylaxis-like reactions, we chose to study two different signal transduction steps, namely calcium influx and cAMP production in mast cells. It is well-recognized that agents that elevate cAMP are effective To further examine the mechanisms by which LA inhibits mast cell-dependent anaphylaxis-like reactions, we chose to study two different signal transduction steps, namely calcium influx and cAMP production in mast cells. It is well-recognized that agents that elevate cAMP are effective inhibitors of mast cell and basophil activity suggesting that this type drug would be beneficial in the treatment of allergic disorders (Weston & Peachell, 1998). Moreover, it has been reported that a rise in cAMP levels can modulate secretory responses by decreasing the free cytosolic calcium concentrations in mast cells (White et al., 1984; Yoshii et al., 1988; Izushi & Tasaka, 1989). In general, stimulation of mast cells with compound 48/80 is believed to initiate the activation of a signal transduction pathway that leads to histamine release. Compound 48/80 initiates generation of superoxide anion by A-kinase inactivation by decreasing the intracellular cAMP concentration in RPMCs (Fukuishi et al., 1997). Generated superoxide anion results in inositol 1,4,5-triphosphate (IP3) or GTP-induced calcium release from the endoplasmic reticulum (ER) (Akagi et al., 1994). Accordingly, depletion of the intracellular calcium store activates a calcium influx into RPMCs, which leads to an increase in intracellular calcium content (Hoth & Penner, 1993). Consequently, increased intracellular calcium levels cause histamine release from RPMCs (Pearce et al., 1981). Interestingly, treatment with LA transiently increases cAMP levels beyond the basal level. Although the mechanism of LA-induced cAMP production has not been elucidated, LA may activate adenylate cyclase directly or indirectly, to otherwise inhibit cAMP phosphodiesterase. In addition, LA prevents compound 48/80-induced calcium uptake into RPMCs in a dose-dependent fashion. Taken together, these results indicate that LA may block the generation of superoxide anion by A-kinase activation by increasing intracellular cAMP in 48/80-treated RPMCs. Hence, we speculate that an LA-induced decrease of superoxide anions as well as an increase of cAMP impedes histamine release by decreasing intracellular calcium content through inhibition of not only IP3 or GTP-induced calcium release from ER but also calcium uptake into compound 48/80-treated RPMCs (Yoshii et al., 1988; Akagi et al., 1994).
However, we cannot firmly rule out the possibility that cAMP is unimportant as an initiator of the secretory response in mast cells. Studies with RPMCs have reported that an increase and decrease in cAMP are not always associated with inhibition and mediation of secretion, respectively (Weston & Peachell, 1998). Moreover, extension of these studies to human cells has revealed that cAMP is not related to the initiation of the secretory process (Peachell et al., 1988; Weston & Peachell, 1998). Therefore, the possibility that the increase of cAMP by LA may not inhibit mediator release cannot be excluded. Although current opinion suggests that cAMP is not important in initiating the secretory response in mast cells, the cAMP signaling system affords targets for pharmacological intervention and any agent that augments cAMP in mast cells could have therapeutic possibilities in allergic conditions such as asthma. Thus, future studies will be required to elucidate the details involved in the relationship between cAMP levels and the secretory response by RPMCs.
In conclusion, the results obtained in the present study provide evidence that LA contributes to the prevention or treatment of mast cell-mediated allergic diseases by inhibiting the release of histamine. The LA-mediated inhibition of mediator release from mast cells may be due to inhibition of calcium uptake and augmentation of intracellular cAMP levels. These notions along with the known anti-oxidant properties of LA suggest that further evaluation of its utility in the treatment of immediate-type allergic diseases is warranted.



 XML Download
XML Download