

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Autism spectrum disorders (ASDs) are neurodevelopmental disorders characterized by repetitive behaviors and impairments in social interactions and language [12]. The results of numerous studies have suggested that genetics and the environment, especially during pregnancy and the fetal period, play pivotal roles in the etiology of ASD.
Valproic acid (VPA), which is used to treat epilepsy and bipolar disorder, has a teratogenic effect on humans [3]. The prevalence of ASDs in the offspring of mothers treated with antiepileptic drugs during pregnancy has suggested that these medications are associated with ASD [4]. Prenatal exposure to VPA increases the risk of ASD phenotype characteristics, such as repetitive behavior, social impairment, neural tube developmental defects, and excitatory/inhibitory synapse imbalance [5678]. Therefore, prenatal exposure to VPA has been extensively studied in rodents as a model of ASD to better understand the neurobiology of ASD and autistic behavior.
Several lines of evidence, including increased serum, plasma, and brain tissue levels of brain-derived neurotrophic factor (BDNF) in children with ASD, have suggested that BDNF is involved in ASD [91011121314]. Similarly, exposure to VPA has been shown to increase BDNF expression in the immature cortical neuron [15] and fetal brain [16]. Because BDNF is widely expressed in the mammalian brain [17] and is critical for brain developmental mechanisms, such as the regulation of neuritogenesis and synapse formation [18], the aberrant hyperactivity of BDNF may play a key role in abnormal development in ASD [19].
Methyl-CpG-binding protein 2 (MeCP2) is a transcription factor that binds to methylated cytosines in CpG islands and that recruits other transcriptional repressors and histone deacetylases that silence target genes [20]. Numerous studies have reported that MeCP2 controls BDNF expression [20212223]. In addition, the similarities in the expression patterns of BDNF and MeCP2 during the prenatal and postnatal periods [24] and in vivo evidence of functional interactions of MeCP2 with BDNF [21] suggest that MeCP2 is crucial for the regulation of BDNF expression in normal brain development and that abnormalities in BDNF regulation contribute to physiological disorders, such as Rett syndrome.
Tissue plasminogen activator (tPA) is a serine protease that induces the mature and active form of BDNF (mBDNF) through the proteolytic cleavage of the precursor isoform of BDNF (pro-BDNF) [25]. tPA has been reported to increase neurite outgrowth through the degradation of the extracellular matrix or induction of mBDNF [26]. Although stimulatory effects of VPA on tPA have been reported in vascular endothelial cells [27], it is unclear whether VPA affects tPA levels in the developing embryonic brain.
Contradictory results of the regulation of BDNF by VPA have been reported. VPA enhances BDNF expression in primary cultured embryonic neural stem cells [28] and primary cultured neurons [2930], whereas VPA-treated mice exhibited decreased BDNF mRNA levels in the somatosensory cortex [31]. Therefore, in this study, we first investigated the levels of BDNF expression in VPA-treated cortical neural progenitor cells (NPCs).
METHODS
Rat primary cortical neural stem cell culture
These animal studies were approved by the Institutional Animal Care and Use Committee of Chung-Ang University (Approval No. 2017-00093). Primary cortical neural stem cells were cultured from the cortices of embryonic-day-14-old Sprague–Dawley (SD) rats. A Pasteur pipette was used to separate the cortices into single cells, which were then incubated in Dulbecco's modified Eagle's medium/F12 (DMEM/F12) that was supplemented with 20 ng/ml of epidermal growth factor and 10 ng/ml of basic fibroblast growth factor in a 5% CO2, 90% N2, and 5% O2 incubator. The culture media were changed every 2 days until the single cells had grown into floating neurospheres. The neurospheres were dissociated into single cells with trypsin ethylenediaminetetraacetic acid and then regrown into neurospheres in epidermal growth factor- and basic fibroblast growth factor-supplemented media. The neurospheres were dissociated again into single cells and plated on poly-L-ornithine-coated plates with DMEM/F12 media containing 2% penicillin/streptomycin without growth factors or serum. The cells were incubated at 37℃ in a humidified atmosphere with 5% CO2, 90% N2, and 5% O2.
Rat primary neuron culture
Primary cortical neurons were prepared from embryonic-day-18-old SD rats. A Pasteur pipette was used to dissociate the cortices into single cells by pipetting the tissue several times. The cells were seeded onto poly-D-lysine-coated plates and incubated in neurobasal medium containing 1% penicillin/streptomycin, 20 µM glutamine, and B27. The cultures were maintained at 37℃ in a humidified 5% CO2 incubator.
Rat primary astrocyte culture
The prefrontal cortices 2-day-old SD rat pups were digested with trypsin for 10 min at 37℃. The triturated cells were cultured in DMEM/F12 containing 10% fetal bovine serum, 100 U/ml of penicillin, and 100 mg/ml of streptomycin for 7 days. The grown cells were washed twice with serum-free media and then detached using 0.25% trypsin with ethylenediaminetetraacetic acid. The cells were plated at low density (5,000 cells/cm2) on well plates, grown for 3 days, and then used for further analysis.
Casein zymography
tPA activity was measured using plasminogen-casein zymography. The culture supernatants were mixed with zymography buffer [200 mM Tris-HCl (pH 6.8), 8% w/v sodium dodecyl sulfate (SDS), 40% glycerol, 0.02% bromophenol blue, and without β-mercaptoethanol] and separated by electrophoresis on 8% polyacrylamide gels containing casein and plasminogen. After the electrophoresis, the gels were washed twice with 2.5% Triton X-100 for 30 min to eliminate the SDS. The gels were then incubated for 12–24 h at 37℃ in reaction buffer [20 mM Tris-HCl (pH 7.6) and 166 mM CaCl2,] for caseinolysis. To reveal the caseinolytic activity, the gels were stained with 0.1% Coomassie brilliant blue R-250 and destained using destaining solution (20% methanol, 10% acetic acid, and 70% deuterium-depleted water). tPA activity was visualized as the light bands that resulted from casein degradation. The caseinolysis band detected at 68 kDa matched the band of purified tPA standard that was present in the same gel. The gel was analyzed using a LAS 3000 detection system (Fujifilm, Minato-ku, Tokyo, Japan), and the band intensities were measured using ImageJ (https://imagej.nih.gov/ij/).
Western blotting
Twenty micrograms of protein were obtained from the cells and mixed with sample buffer [120 mM Tris-HCl (pH 6.8), 0.5% sodium deoxycholate, 0.1% SDS, and 1% Triton X-100]. The proteins were separated by 8% SDS-polyacrylamide gel electrophoresis and electrically transferred onto nitrocellulose membranes. The membranes were blocked with 5% skim milk at room temperature for 60 min and then incubated with the primary antibodies at 4℃ overnight. The membranes were washed three times with TBS-Tween (0.1%) for 10 min and subsequently incubated with horseradish peroxidase-conjugated secondary antibody for 90 min at room temperature. Each band of interest was detected with enhanced chemiluminescence solution, and the images were captured using LAS-3000. The band intensity was determined using ImageJ software.
Reverse transcriptase-polymerase chain reaction (RT-PCR)
The levels of expression of tPA, MeCP2, and BDNF mRNA were determined by quantitative RT-PCR. The cells were washed twice with ice-cold phosphate-buffered saline (PBS), and total RNA was extracted using TRIzol reagent. First-strand cDNA synthesis was conducted using 2.5 µg of total RNA and reverse transcriptase. The extracted RNAs were incubated at 60℃ for 60 min and then heated at 94℃ for 5 min to terminate the reaction. A total of 5 ng of cDNA was used as a template for the PCR amplification of tPA, MeCP2, and BDNF. The primer sequences were as follows: tPA forward: AGT TGC AGC GAA CCA AGA TG, tPA reverse: TGC CAC GGT AAG TCA CAC CT; MeCP2 forward: CTG CTG CAG AGG CCA AAA AG, MeCP2 reverse: TGG TGG TGA TGA TGG TGC TC; and BDNF forward: GCT GCC TTG ATG TTT ACT TTG, BDNF reverse: ATG GGA TTA CAC TTG GTC TCG T. The PCR products were subjected to 1.0% agarose gel electrophoresis and stained with ethidium bromide. The intensity of each band was analyzed using ImageJ.
Immunofluorescent staining
The cells on the cover glass were fixed with 4% paraformaldehyde at room temperature for 20 min and then washed with PBS. The cells were permeabilized with 0.1% Triton in PBS for 15 min at room temperature and then washed twice with PBS. The cells were incubated with primary antibodies that were diluted with blocking solution (1% bovine serum albumin and 3% fetal bovine serum in PBS) at 4℃ overnight. After washing the cells three times with PBS, they were incubated with secondary antibodies conjugated with Alexa Fluor® 594 Dye and diluted in blocking buffer for 1 h at room temperature. After three washes, diamidino-2-phenylindole (DAPI) that was diluted in PBS was applied to the cells for 5 min. Cover glasses were mounted using Vectashield (Vector Laboratories, CA, US), and the cells were observed using confocal microscopy.
Statistical analysis
Data were displayed as mean±standard error of mean (S.E.M.) and were analyzed for statistical significance using one-way analysis of variance (ANOVA) followed by post hoc Tukey's comparisons test. Differences were considered statistically significant when the p value was less than 0.05 (p<0.05). All statistical analyses were conducted using GraphPad Prism software.
RESULTS
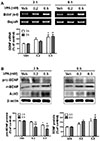
VPA increased BDNF expression in cultured cortical NPCs
To investigate whether VPA administration affected BDNF in developing brain, we first examined BDNF expression in VPA-treated embryonic cortical NPCs. Of the 9 exons, only exons 1, 4, and 6 of BDNF were detected in the fetal mice brains, and the levels of these exons were increased by VPA treatment [16]. However, only BDNF exon 4 expression was significantly increased by VPA treatment in the prefrontal cortex [32]. Thus, we then measured the expression levels of BDNF exon 4 after treatment with VPA (0.2 and 0.5 mM) in NPCs. VPA increased the expression of BDNF exon 4 in a concentration-dependent manner (Fig. 1A). To further investigate this, we investigated the levels of expression of pro-BDNF and mBDNF in VPA-treated NPCs. The expressions of mBDNF and acetylated histone 3 were increased, whereas pro-BDNF expression levels were suppressed by VPA (Fig. 1B). Therefore, these results suggested that VPA increased BDNF transcription in NPCs.
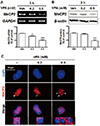
VPA decreased MeCP2 expression in NPC nuclei
Because BDNF exon 4 is the preferential target for epigenetic regulation as it contains the specific binding site for MeCP2, the epigenetic regulation of BDNF is thought to be involved in physiological conditions, such as psychiatric disorders [33]. MeCP2 suppresses BDNF gene transcription [3435] by binding to methylated DNA in promoter 4 of Bdnf, which maintains the repressed state of the Bdnf gene [23]. Therefore, we investigated the effects of VPA on MeCP2 expression in VPA-treated NPCs. The levels of MeCP2 mRNA and protein were significantly decreased by treatment with 0.5 mM of VPA (Figs. 2A and B). In addition, the immunostaining results showed that MeCP2 expression was decreased by VPA treatment (Fig. 2C). Interestingly, the diffused pattern of MeCP2 expression was decreased by VPA treatment, which resulted in the dissociation of MeCP2 from the DAPI puncta region that contains enriched A/T bases.
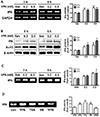
VPA increased the levels of tPA expression and activity in cultured NPCs
Our results showed that VPA increased mBDNF levels and suppressed pro-BDNF levels, which suggested that the processing of pro-BDNF into mBDNF was enhanced. Therefore, we investigated the levels of tPA expression in VPA-treated NPCs. The mRNA and protein levels of expression of tPA and acetylated histone 3 were increased in NPCs by VPA treatment (Figs. 3A and B). In addition, the caseinolytic assay showed that tPA activity was enhanced by VPA (Fig. 3C). To investigate whether VPA upregulated tPA through histone regulation, we treated the NPCs with other HDACis. TSA and SB also enhanced tPA activity, while valpromide, a VPA analog that lacks histone deacetylase inhibition activity [3637], failed to enhance the activity (Fig. 3D). These results indicated that VPA increased tPA expression in NPCs through histone modification.
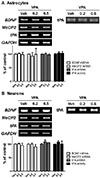
VPA did not affect MeCP2 and tPA expression in neurons and astrocytes
VPA has previously been shown to increase BDNF expression in astrocytes [38], astrocytes from LEW/N rats [39], and human astrocytoma cells [18]. Moreover, a protective role of VPA was suggested by the increased expression of BDNF in neurons [1540] and neuroblastomas [41]. To investigate whether VPA increased BDNF in other neuronal cells through the regulation of MeCP2 and tPA, we treated neurons and astrocytes with VPA. VPA treatment did not result in significant differences in the levels of BDNF, MeCP2, and tPA expression or tPA activity (Figs. 4A and B). Therefore, these results suggested that the VPA-induced stimulation of BDNF in cortical NPCs occurred through the regulation of MeCP2 and tPA expression.
DISCUSSION
In the present study, we found that VPA upregulated the levels of mBDNF in NPCs through dual mechanisms involving enhanced BDNF transcription and maturation. We found that the mRNA and protein expression levels of BDNF were increased, whereas those of MeCP2 were decreased, by VPA treatment. In addition, the levels of mRNA, protein, and activity of tPA were enhanced in VPA-treated NPCs. In addition, other histone acetylation inhibitors (HDACi) such as trichostatin A (TSA) and sodium butyrate (SB) enhanced tPA activity, while valpromide, which is an analog of VPA that lacks HDACi activity, failed to induce tPA activity. These results indicated that VPA stimulates BDNF expression in NPCs through the regulation of MeCP2 and tPA, which may contribute to aberrant cortical development in ASD.
Numerous studies have reported that stimulated BDNF is involved in the abnormal development of the developing brain. Because BDNF levels are normally low in the fetal brain and gradually increase during maturation [42], transient increases in BDNF levels might play a causal role in embryonic brain development defects, such as altered neuronal laminar fate and premature exit from the cell cycle in cortical development [43]. A rapid and uncorrected increase in BDNF levels by VPA treatment would leave an “architectural signature” during embryonic cortical development [44] and might lead to permanent defects in the postnatal period. Interestingly, VPA rapidly stimulated BDNF mRNA expression in 3 or 6 h, which was also observed in VPA-exposed embryonic-day-12.5-old fetal brain [16], whereas the VPA enhancement of BDNF expression took one [15] or a few days [2829]. In addition, postmortem brain studies have shown increased immunoreactivity of BDNF in patients with ASD due to the suppression of BDNF cleavage, which was independent of transcriptional regulation [13].
In present study, VPA-induced upregulation of BDNF was observed only in NPCs not in astrocytes or neurons. In other studies, the upregulation of BDNF by VPA was investigated at relatively later time point after VPA exposure, from 24 h to a few days in neurons or astrocytes [303245]. In addition, the concentration range of VPA was 0.6–1.2 mM [3846], due to 0.3 and 0.4 mM of VPA did not have significant effect in astrocytes [46]. In present study, we investigated the effect of VPA on BDNF expression at relatively in earlier time, 3 and 6 h, and VPA concentration was lower (0.2 and 0.5 mM). Therefore, it assumes that early exposure and lower concentration of VPA would be a causal factor of no significant effect of VPA in BDNF expression in astrocytes or neurons. VPA would promote BDNF expression at higher concentration of VPA and further exposure time in astrocytes or neurons.
In addition, cell-type specific acetylation and methylation status would be a causal role of VPA-induced regulation of BDNF in NPCs. In recent study, cell-type specific ChIP-seq analyses displayed histone modification, such as acetylation and methylation of histone were variable in cell-types, which determine cell-type specific transcripts levels [47]. In addition, histone acetylation levels are gradually decline during differentiation of embryonic stem cells [48]. Considering VPA enhanced BDNF protein only in E12.5 fetal brain but not in matured brain [16], the developing brain seems to be more sensitive to histone acetylation than the matured brain. Although regional differences of histone acetylation have been observed during developing mouse embryonic brain and chick spinal cord [49], further studies would be required to clarify the cellular difference in acetylation and methylation status which determine cell-type specific transcripts levels in response to histone modification.
Because BDNF can cross blood brain barrier [5051], highly correlation has been suggested between plasma and cortical BDNF levels [5052]. It has been reported BDNF level in serum was significantly decreased in bipolar patients compared to healthy controls [535455], and the enhanced BDNF was observed in patients treated with antidepressants [535657]. Likewise, upregulated BDNF was monitored in bipolar patients treated with VPA and dextromethorphan [54], suggesting VPA have therapeutic effects on bipolar patients by promoting BDNF level. However, in other studies, serum BDNF level in bipolar patients was not significantly different compared to healthy control [58]. In addition, the BDNF levels in depressed patients treated with antidepressants were constitutively suppressed compared to healthy control [59], and serum BDNF was independent with antidepressant treatment [60], suggesting correlation between level of BDNF and VPA is required to be further investigated.
The results of this study showed that the VPA increase of BDNF mRNA expression accompanied the suppression of the expression of MeCP2, which is a transcriptional repressor of BDNF expression. The dysfunction of MeCP2 is related to Rett syndrome [6162], and Rett-like features are induced by MeCP2 inactivation in adult brain [63]. A regulatory role of MeCP2 on BDNF expression has been suggested by three different models, including repression, activation, and dual operation [24]. Interestingly, the intracellular levels of BDNF are enhanced by both the knockdown as well as the overexpression of MeCP2 in hippocampal neurons [64], which suggests that adequate levels of MeCP2 are required for defining the optimal levels of BDNF. MeCP2 binds at the methylated CpG region that contains enriched A/T bases in Bdnf genes [65]. Our immunostaining results that showed that the nuclear expression of MeCP2 was mainly decreased in the DAPI puncta region suggested that VPA preferentially decreased the levels of MeCP2 expression in the A/T region.
Accumulating evidences demonstrated that microRNA plays crucial role in neural development [6667], and perturbation of microRNA is involved in neurodevelopmental disorders, such as ASD and fragile X-syndrome [686970]. MiR-132 is one of the dominantly expressed microRNA in brain [71], and it represses MeCP2 levels by binding its miRNA recognition element (MRE) with MeCP2 3′UTR thus inhibits MeCP2 levels [72]. Interestingly, prenatal exposure to VPA displayed enhanced miR-132 whereas decrease of MeCP2 mRNA in embryonic brain [71]. Likewise, upregulated miR-132 in contrast to suppressed MeCP2 was observed in TSA treated NIH/3T3 cells [73], suggesting it would be possible VPA regulates MeCP2 expression through regulation of miR-132 levels.
In this study, we found that VPA enhanced the expression of tPA in NPCs. VPA also enhances tPA expression in cultured endothelial cells [27], and enhanced vascular tPA production decreases in vivo fibrin accumulation [74]. Reduced incidences of strokes and myocardial infarctions have been observed in patients with epilepsy who were treated with VPA [7576], which also suggests that VPA enhances tPA activity. tPA expression is enhanced by other HDACis, such as TSA, SB, and MS-275, in cultured human endothelial cells [777879], and tPA activity is not altered in the absence HDAC regulation; this suggests that histone acetylation is required for the upregulation of tPA [27]. In those study, chromatin immunoprecipitation assay displayed acetylated histone was enhanced in tPA transcription start site by VPA. The marginal enhancement of mRNA for PAI-1 or uPA, the other plasminogen activator [27], suggesting tPA is sensitive to histone acetylation. Indeed, tPA promoter contains binding sites for transcription factor Sp1, which is induced by HDAC inhibitors [808182], and that is critical for consecutive expression of tPA [8384]. Therefore, it assumes that HDACi promotes tPA transcription by inducing Sp1 expression in NPCs.
VPA has been reported to promote neurite outgrowth in NPCs [28]. Because tPA promotes neuritogenesis through the phosphorylation of LRP5/6 [85], tPA might play a causal role in the enhanced neurite outgrowth observed in VPA-treated NPCs. Although VPA did not directly stimulate tPA expression, VPA promoted tPA activity by suppressing the expression of the tPA inhibitor plasminogen activator inhibitor-1 (PAI-1) [86]. PAI-1 is highly expressed in astrocytes [87], and it terminates tPA activity by forming a complex with the catalytic domain of tPA [88]. We previously showed that tPA, but not PAI-1, is expressed in NPCs [85]. These findings suggest that the rapid induction of tPA by VPA resulted from the promotion of tPA expression by VPA, which was independent of PAI-1 activity.
As several studies have reported, a number of genes play a role in the abnormal brain and synapse development of subjects with ASD [8990]. Likewise, our study showed that the exposure to VPA increased the risk of ASD through changes in the regulation of the expression of multiple genes in NPCs. In the future, studies should investigate the contribution of the rapid modulation of gene expression by histone remodeling towards abnormal gene expression in ASD development.
 XML Download
XML Download