

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Cancer is one of the leading causes of death worldwide. Cancer cells undergo rapid, uncontrolled proliferation, which makes the microenvironment to hypoxic condition, results in depleted the oxygen, and insufficient blood supply. To fulfill their demands for nutrient, oxygen, and other growth factors, angiogenesis is required. Several studies have revealed that targeting angiogenesis is an effective approach for cancer therapy under hypoxic conditions [12]. However, the molecular mechanisms underlying this process remains unclear.
Endothelial progenitor cells (EPCs) were first discovered and isolated by Asahara et al. in 1997 [3]. They found that EPCs have the ability to form new blood vessels in a process known as neovascularization. EPCs can be isolated from CD34+ cells in human cord blood, peripheral blood, as well as the bone marrow. They express the surface markers for CD34, CD31, and vascular endothelial growth factor receptor-2 (VEGFR-2, KDR) [45]. It has been established that EPCs contribute to tumor development and metastasis via tumor angiogenesis [6]. In addition, they demonstrate the ability to differentiate into endothelial cells (EC), which is stimulated by pathological conditions such as hypoxia. Hypoxia-induced angiogenesis is consist of several successional steps including 1) angiogenic factors (such as VEGF, bFGF) released from hypoxic cells, and bind to EC / EPC, 2) activate MMPs to degradate the extracellular membrane (ECM), then 3) cell migration and invasion, 4) tube formation, 5) vessel stabilization, in briefly. Therefore, understanding the mechanism of hypoxia-induced angiogenesis and finding a new approach to prevent angiogenic function are essential for development of cancer therapy.
Recently, mitochondrial dynamics has emerged as a critical mechanism for cellular function in hypoxic cell signaling. Mitochondria are dynamic subcellular organelles that undergo continuous fission and fusion to maintain mitochondrial function and energy metabolism [78]. These processes are tightly regulated by specific molecules, such as dynamin-related protein Drp1 for fission, and MFN1/2 and OPA1 for fusion. Homeostasis between mitochondrial fission and fusion is important for sustaining cell survival, proliferation, and differentiation. For example, mitochondrial fission induced by CDK1/cyclinB/pDRP1 axis is essential for cell mitosis [9], and phosphorylation of DRP1 at Ser637 induced by nutrient starvation sustain the cell survival via inhibition of mitochondrial fission [10]. Kasturi et al. reported that DRP1-dependent mitochondrial fission is essential for cell differentiation [11], Luchsinger et al. showed that MFN2-induced mitochondrial fusion plays critical role to maintain stem cells [12]. It has been well-established that a shift towards mitochondrial fission occurs under hypoxic conditions [1314]. Most of the studies to date have focused on the relationship between mitochondrial fission, increased ROS production, and apoptosis [1516]. A few reports have also investigated the effect of accelerated mitochondrial fission on cell migration and invasion in breast cancer [17]. However, little is known about the effect of mitochondrial fission on endothelial cells during angiogenesis.
Therefore, the aim of this study is to clarify the role of mitochondrial fission in hypoxia-induced angiogenesis. This study demonstrated that specific inhibition of DRP1-mediated mitochondrial fission attenuated the EPC bioactivities under hypoxia. We suggest the regulation of mitochondrial dynamics maybe a novel therapeutic approach for cancer therapy.
METHODS
Isolation of endothelial progenitor cells
Human EPCs were isolated as previously described [18]. All experimental procedures have been reviewed and approved by the International Review Board (IRB) of Pusan National University Hospital (IRB No.05-2017-053). In brief, mononuclear cells (MNCs) were separated from human umbilical cord blood using Ficoll-Paque PLUS (GE Healthcare, Uppsala, Sweden), according to manufacturer's instructions. MNCs were cultured with the EGM-2 Bullet Kit system (Lonza, Walkersville, MD, USA) for five days in 1% gelatin-coated plates. Cultures were supplemented with 1X penicillin streptomycin (Welgene, Korea). EPCs formed spindle-shaped colonies. To generate hypoxic conditions, cells were starved with 1% FBS in EBM-2 media for 12 h, followed by a 24-h culture period at 1% O2.
Cell viability assay
Mdivi-1 was obtained from Sigma Aldrich, and was dissolved in DMSO to yield a stock concentration of 5 mM. To assess cell viability, 1×105 EPCs were seeded in 96-well plates for 24 h. Following stabilization, EPCs were treated with serially diluted Mdivi-1 (0, 3.125, 6.25, 12.5, 25, 50, 100 µM) for 24 h. The effect of Mdivi-1 on cell viability was measured using D-Plus CCK cell viability assay kit (Dongin LS, Korea), according to manufacturer's protocol. To quantify cell viability, absorbance was measured at 450 nm with a TECAN microplate reader; results were normalized and compared with those of the untreated control.
siRNA-mediated knockdown of DRP1 gene expression
DRP1 siRNA (siDRP1) was a kind gift from Prof. Young chan Chae (UNIST), and scramble siRNA (scRNA) was purchased from Dharmacon (USA). Cells were seeded into 1% gelatin-coated 6-well plate and induced the transfection with either siDRP1 or scRNA using Lipofectamine RNAiMAX reagent (Thermo Fisher Scientific Inc.) following manufacturer's instruction. 6 h after transfection, media was replaced with EGM-2 culture media. Further experiment was conducted 24–48 h after transfection.
Western blot
Human EPCs were lysed with Pro-prep (iNtRON biotechnology, Korea). Whole cell lysates (25 µg) were separated by either 8% or 15% SDS-PAGE, after which membranes were blocked with 5% skim milk for 1 h at 25℃. Membranes were probed with specific antibodies, and proteins of interest were visualized with HRP-conjugated secondary antibodies (Thermo Fisher Scientific Inc., MA, USA) and Luminata Crescendo HRP substrates (Millipore, MA, USA). Primary antibodies against mouse anti-HIF-1A (1:1000), mouse anti-DLP1 (1:1000, all above from BD Biosciences, NJ, USA), rabbit anti-phospho-DRP1 (Ser616) (1:1000), rabbit anti-phospho-DRP1 (Ser637) (1:1000, Cell Signaling Technology, Inc., MA, USA), rabbit anti-Fis1 (1:1000), rabbit anti-OPA1 (1:1000, all above Abcam, Cambridge, UK), rabbit anti-MFN1 (1:1000), and mouse anti-beta-actin (1:2500, all above Santa Cruz Biotechnology, CA, USA) were used.
Cell migration and invasion
Human EPC migration assays was performed using 8 µm-pore transwell inserts (Corning Inc., MA, USA). For migration assays, 2×105/ml cells were suspended in EBM-2 medium; 100 µl (20 000 cells) cell suspension was added into the upper chamber, and 500 µl EGM-2 media was placed in the lower chamber. For invasion assays, 50000 cells were seeded in Matrigel-coated transwell inserts (BD Biosciences, NJ, USA). After 24 h of incubation under normal or hypoxic conditions (1% O2), cells were fixed with 4% paraformaldehyde for 10 min, followed by staining with 0.5% crystal violet in 25% methanol for 30 min at 25℃. Cell migration and invasion were visualized with a Zeiss Axio Imager M2 optical microscope (Zeiss, Munchi, Germany), and cell counting was conducted with the Image J software.
Tube formation
Matrigel growth factor reduced (GFR) Basement Membrane Matrix (100 µl) (Corning Inc., MA, USA) was placed into 96-well plates, and was incubated at 37℃ and 5% CO2 for 1 h. Then, 1×104 cells were seeded under normal or hypoxic conditions. Tube formation was observed for 6–18 h. In inhibition experiments, cells were pre-treated with 12.5 µM Mdivi-1 for 24 h prior to tube formation. Total tube length was quantified using the Image J software.
FACS analysis
Cells were suspended in FACS buffer (1% FBS, 2 mM EDTA in PBS) and stained with specific antibodies including FITC-conjugated anti-CD34, APC-conjugated CXCR, Alexa Fluor 647-conjugated CD31, PE-conjugated KDR (all from BD Biosciences, NJ, USA), and APC-conjugated c-kit (Miltenyi Biotec, Germany). All antibodies were diluted 1:100; cells were incubated in primary antibodies for 30 min at 4℃. After cells were washed twice with cold PBS, they were resuspended and analyzed with flow cytometry.
Immunofluorescence and confocal microscopy
Human EPCs were seeded at a density of 4×104 cells/ml in 1% gelatin-coated cover glass. After 24 h, untreated or Mdivi-1 treated cells were cultured in either normal or 1% O2 for 24 h. Cells were then fixed with 4% paraformaldehyde for 15 min, and were permeabilized with 0.25% PBST for 10 min at room temperature. Unspecific binding was blocked (1% BSA in PBST) for 1 h at room temperature, followed by staining with mouse anti-Tom20 antibody (1:100, Santa Cruz, CA, USA) at 4℃ overnight.
Statistical analysis
All data were presented as mean±standard error of mean (SEM). Descriptive statistics were analyzed using the GraphPad Prism 5 software (GraphPad Software Inc., La Jolla, CA). We used Student's unpaired t-tests for comparison between two groups. p<0.05 was the accepted value for statistical significance.
RESULTS
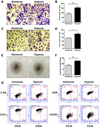
Hypoxia induces EPC bioactivities
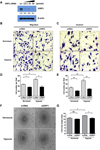
To confirm the effect of hypoxia on EPCs bioactivities, we performed angiogenic functional studies such as migration, invasion, and tube formation assays. Results indicated that hypoxia increase cell migration and invasion by 1.7- and 1.4-fold, respectively (Figs. 1A-D). Similarly, cells under hypoxic conditions showed greater ability to form tubes as compared to that of control, which is an indicator of endothelial cell differentiation (Figs. 1E and F). To assess whether hypoxia increases EPC population, we evaluated the expression of EPC surface markers such as CD31, KDR (VEGFR), c-kit, CXCR4, and CD34 using flow cytometry. No significant change was observed between normal and hypoxic conditions. Cells that were double positive for CXCR4, a known for migration marker, and CD34, a marker of EPC, were increased approximately by 1.7-fold under hypoxic conditions (Fig. 1G). However, other markers such as c-kit, a known for stemness, and CD31, a marker of endothelial lineage, and KDR, a one of the marker of EPC, showed slightly change. Taken together, our results showed that hypoxia increases cell migration, invasion, and tube formation, but does not have an effect on the EPC population.
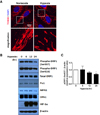
Mitochondrial fission-related dynamics in EPCs is induced in response to hypoxic signals
Recently, it has been reported that mitochondrial dynamics is one of the essential mechanisms of hypoxic cell signaling. To verify the effect of hypoxia on the mitochondrial dynamics in EPCs, we firstly detect the mitochondrial dynamics using confocal microscopy. We observed that mitochondrial fission was induced under hypoxic condition (Fig. 2A). In addition, the expression of mitochondrial fission/fusion-related gene was changed (Fig. 2B). DRP1 is a key regulator of fission, and phosphorylation of DRP1 at Ser616 is typically used as an active form of DRP1. Dephosphorylation at Ser637 on DRP1 is also known to accelerate fission. Based on our results, the expression of pDRP1 (Ser637), an inhibition marker of fission, was significantly decreased in under hypoxic conditions at time point 24 h (Fig. 2C). Unexpectedly, pDRP1 (Ser616) was decreased during hypoxia. Fis1 is an adaptor of DRP1, which retains DRP1 in the mitochondria to accelerate fission. We observed that 24 h post-hypoxia, the expression of Fis1 was slightly increased, similar pattern to pDRP1 (Ser637). The expression of MFN1, OPA1, the markers of mitochondrial fusion, were also decreased. Taken together, we suggest that pDRP1 (Ser637)/Fis1/MFN1/OPA1 orchestrate inducing mitochondrial fission under hypoxic condition.
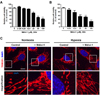
Specific inhibition of DRP1 blocks hypoxia-induced mitochondrial fission in EPCs
To confirm DRP1 is involved in hypoxia-induced mitochondrial fission, we used Mdivi-1, a known specific inhibitor of DRP1. First, we determined the optimal concentration of Mdivi-1 using the CCK-8 cell viability assay. At concentrations above 25 µM, EPCs showed cytotoxicity and reduced cell viability. Therefore, the concentration of Mdivi-1 for our 24-h incubation experiments was set at 12.5 µM (Figs. 3A and B) without cell toxicity. As expected, confocal microscopy data demonstrated that hypoxia-induced mitochondrial fission was blocked by Mdivi-1 treatment (Fig. 3C). Therefore, we confirmed that hypoxia-induced mitochondrial fission is mediated by DRP1 in EPCs.
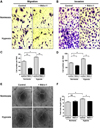
Specific inhibition of DRP1 reduces hypoxia-induced EPC bioactivities
To validate the role of DRP1-mediated mitochondrial fission in hypoxia-induced EPCs bioactivities, we assessed migration, invasion, and tube formation using DRP1 siRNA and DRP1 specific inhibitor. In the presence of Mdivi-1, hypoxia-induced cell migration was blocked by 2.8-fold change, and cell invasion also showed a trend for reduction with Mdivi-1 treatment (Figs. 4A-D). In addition, the ability for tube formation was reduced during DRP1 inhibition under hypoxic conditions (Figs. 4E and F). To confirm the role of DRP1-mediated mitochondrial fission in EPC angiogenic function, we performed the DRP1 siRNA experiments. We first confirmed that DRP1 expression was dramatically decreased up to 90% (Fig. 5A). DRP1 silencing showed decreased cell migration in 1.7-fold change, and decreased in cell invasion by 2.1-fold change under hypoxia (Figs. 5B-E), similar pattern to Mdivi-1 treatment. The ability of tube formation also decreased in DRP1 silencing cells, therefore we demonstrated that DRP1-mediated mitochondrial fission is essential step for hypoxia-induced EPCs angiogenesis. As expected, no significant changes were observed in EPC surface markers with Mdivi-1 treatment (Fig. 6). Collectively, these data indicated that DRP1 is essential in hypoxia-induced EPC migration, invasion, and tube formation, but does not play a role in expression of EPC surface markers.
DISCUSSION
In this study, we demonstrated that key role of DRP1-mediated mitochondrial fission in hypoxia-induced EPCs bioactivities; this is supported by data from previous studies. We observed that hypoxia-induced mitochondrial fission occurs in EPCs, which is correlated with evidence from previous reports in neuronal cells, cardiomyocytes, and cancer cells [171920]. In addition, phosphorylated DRP1-Ser637, a suppression marker for mitochondrial fission, was impaired in hypoxic conditions. This suggested that pDRP1-Ser637 is a key molecule that mediates hypoxia-induced mitochondrial dynamics in EPCs. Coordinately, the expression of MFN1, OPA1 also showed reduction, and Fis1 tends to slightly increase under hypoxic conditions. Finally, targeting for DRP1 using Mdivi-1, a selective DRP1 inhibitor, and DRP1 siRNA, showed dramatically impaired EPC migration, invasion, and tube formation even under hypoxic conditions. Therefore, we suggest that modulation of mitochondrial fission could be key mechanism for hypoxia-induced angiogenesis.
It is the first time to discover the role of hypoxia-induced mitochondrial fission in EPC-mediated angiogenesis. While several studies have reported that ischemia reperfusion-induced mitochondrial fission is associated with decreased ROS production and apoptosis in neuronal cells and cardiomyocyte [151621], little is known about the effect of mitochondrial dynamics on angiogenesis. As hypoxia is known to affect both angiogenesis and mitochondrial function, we examined that the effect of mitochondrial dynamics on angiogenesis [22]. In agreement with our results, Zhao et al. confirmed that DRP1 plays critical roles in cell migration and invasion in breast cancer under hypoxic condition [23]. They also observed reduced cell migration and invasion under Mdivi-1 treatment, similar pattern to our result (Fig. 4A). In endothelial cells (ECs), little is known about the relationship between mitochondrial dynamics and cellular function. Lin et al. reported that decreased DRP1 and mitochondrial fusion is involved in senescence, but not in hypoxia-induced angiogenesis [24]. They demonstrated that Mdivi-1 treatment decreases cell proliferation, oxygen consumption rate (OCR), and G2/M cell cycle arrest via autophagic flux. However, the critical regulatory site for DRP1 phosphorylation during senescence remains unclear. In addition, Kashatus et al. reported that MAPK/Erk2/pDRP1 axis is critical for tumor growth [25]. They showed that targeting DRP1 impaired tumor growth, and that phosphorylation of DRP1 at Ser616 is critical for tumor development. Therefore, these reports support our hypothesis that targeting of mitochondrial dynamics is an effective strategy that not only inhibits tumor growth, but also impairs angiogenesis at tumor sites.
In our results, pDRP1 (Ser637) seems to be a key regulatory site rather than pDRP1 (Ser616). Unexpectedly, the expression of pDRP1 (Ser616) was slightly but not significantly decreased in hypoxia. The expression of MFN1, and OPA1 tends to decrease in hypoxia. We also observed the mitochondrial fission using confocal microscopy (Fig. 2A). Taken together, pDRP1 (Ser637), Fis1, MFN1, and OPA1 coordinately induce the mitochondrial fission during hypoxic condition, except pDRP1 (Ser616). Maybe there are two possible reasons to describe the expression of pDRP1-Ser616 in hypoxic condition. First, decreased expression of pDRP1-Ser637 maybe major factor to induce mitochondrial fission rather than pDRP1-Ser616 in EPC. The response to hypoxia maybe different dependent on cell types, so pDRP1-Ser637/OPA1/MFN1 orchestrate inducing mitochondrial fission especially in EPC. Second, hypoxia-induced phosphorylation site of DRP1 is controversial, yet. Because upstream signaling of pDRP1-Ser616 and pDRP1-Ser637 are different and complex. Kim et al. reported that decreased pDRP1-Ser637 induced by AKAP121/Siah2 axis plays essential role for mitochondrial fission to adapt the hypoxic condition [26], whereas Marsboom et al. reported that HIF1A/cyclin B1/CDK1 induce DRP1 phosphorylation at Ser616, which leads to mitochondrial fission and pulmonary arterial smooth muscle proliferation [27]. Taken together, our results showed that pDRP1-Ser637, OPA1, MFN1 induce the mitochondrial fission response to hypoxic condition, especially in EPCs.
However, there is a missing gap between mitochondrial dynamics and angiogenesis. To address the missing gap, there are several possible hypotheses such as energy depletion, autophagic flux, and subcellular redistribution. According to Zhao et al., they addressed the mitochondrial fission is benefit to redistribution of mitochondrial to lamellipodia, which is essential step for cell migration [23], whereas mitochondrial fusion block that process. Another hypothesis is mitochondrial fission-induced energy depletion leads to inhibit the cell migration and invasion. Silvia et al. reported that oligomycin A, an inhibitor of mitochondrial F1F0-ATPase, inhibit the cell migration, and invasion because of lack of energy source, even under the DRP1-induced mitochondrial fission [28]. The other possible hypothesis is mitochondrial fission is involved in autophagic flux. Mdivi-1 treatment induced premature senescence and impaired angiogenic function by suppressing autophagic flux. In contrast aberrant DRP1 expression restored autophagic flux and angiogenic function [24]. To address the precise mechanism of mitochondrial fission in EPC-mediated angiogenesis, further experiment is required.
Mdivi-1, a selective DRP1 inhibitor, has been reported to be a potential therapeutic target for cancer due to its anti-cancer effect [29]. Previous studies generally used 50 µM Mdivi-1 for treatment in cancer cells (IC50=50 µM) [30]. However, the sensitivity of EPCs to Mdivi-1 was four times higher as compared with that of cancer cells (Figs. 3A and B). According to our results, a dose of 12.5 µM Mdivi-1 exerted no significant effect on EPC viability, with or without serum starvation. To minimize cell toxicity, 12.5 µM Mdivi-1 was used for further studies. We investigated the effect of Mdivi-1 on EPC bioactivities, namely, cell migration, invasion, and tube formation under hypoxic condition. Mdivi-1 treatment showed significantly decreased migration, invasion, and tube formation under hypoxia condition. That results showed similar pattern to DRP1 silencing experiment. Therefore, we suggest that targeting of DRP1 could be effective in preventing angiogenesis. However, we did not observe any significant effect of Mdivi-1 on EPC surface marker expression. While a slight change was detected, it was not sufficient to affect cell migration, invasion, and tube formation. Therefore, we confirmed that regulation of mitochondrial fission-fusion affects EPC bioactivities, but not the population of the EPC.
Taken together, we have presented a new perspective in cancer therapy, and have demonstrated that mitochondrial dynamics plays an important role in tumor angiogenesis. Furthermore, we suggest that specific modulation of mitochondrial dynamics under hypoxic conditions can be used as a potential therapeutic strategy in tumor angiogenesis.

 XML Download
XML Download