

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Allergic diseases including food allergies, conjunctivitis, allergic asthma and rhinitis, are on a rise worldwide, affecting approximately 1 out of 5 people [1]. Bronchial asthma is a common allergic disease with hallmark symptoms including inflammation of the airways, airway hyperresponsiveness (AHR), cough, dyspnoea, wheezing mucus hypersecretion, and airway remodelling [23]. The T helper 2 (Th2) cells are crucially involved in allergic asthma. Th2 cells produce cytokines—interleukin (IL) 4, IL-5, and IL-13 that are majorly responsible for inflammation in asthma [45]. IL-4 and -13 induce the production of immunoglobulin E (IgE) while IL-5 exerts vital roles in differentiation and activation of eosinophils [67]. Immune responses exerted by Th2 cells is balanced by Th1 cells that secrete interferon (IFN)-γ. IL-4 is involved in class switching of IgG1 and in synthesis of IgE, while, IFN-γ is implicated in the class switching of IgG2α levels of allergen specific IgE and Th1/Th2 cytokine balance is vital in asthma assessment [8]. Th17 cells, another subset of T helper cell secretes several cytokines, including IL-17, IL-6, IL-22, and TNF-α [910]. IL-17 is associated with recruitment for airway macrophages [11].
Janus kinase-signal transduction and activation of transcription (JAK/STAT) pathway, a major cell signaling cascade, is well documented in Th2 cell differentiation [12] and in Th17 cell differentiation, survival and proliferation [13]. JAKs are regulators of signaling via the cytokine receptors. JAK1 and JAK3 play crucial roles in the initiation of inflammatory responses [14]. Studies have reported that the JAK-STAT pathway in asthma [15] and in allergic rhinitis [16]. It is known that allergic diseases are predominantly characterized by elevated levels of IL-4, IL-5, IL-13 and IL-31. Further, binding of these cytokines with their respective receptors activate the JAK/STAT signaling [17].
Further, Notch signaling pathway is also reported to be involved in pathogenesis of asthma [18]. Notch1 is found to play a distinct role in T cell development and in Th1/Th2 imbalance in asthma [1819]. Studies have shown increased protein levels of Notch1 in ovalbumin (OVA)-sensitized mice [2021]. Also, inhibition of Notch1 signaling by γ-secretase inhibitor was found to cause a decrease in Th2 cytokine production while it raised Th1 cytokine levels, thus aiding in alleviation of asthma-induced pathological changes [20].
In recent years, a number of plant-derived compounds have been found to regulate JAK/STAT pathway and inflammatory responses [32223]. Magnolol, a neolignan isolated from Magnolia officinalis, is found to exert a broad spectrum of pharmacological effects including anticancer [2425], antioxidant and anti-inflammatory activities [2627]. The present study we explored the effects of magnolol on inflammatory responses and on JAK/STAT and Notch signaling in OVA-sensitized mice model of asthma.
METHODS
Chemicals and antibodies
Antibodies against JAK1, JAK2, JAK3, STAT1, STAT3, STAT6, p-STAT1, p-STAT3, p-STAT6, β-actin (Cell Signaling Technology, Beverly, MA, USA), Jagged1, Jagged2 and Notch1 (Santa Cruz Biotechnology, Santa Cruz, CA, USA) were used for expression analysis. OVA, magnolol, methacholine and other chemicals and reagents used were got from Sigma-Aldrich (St. Louis, MO, USA) otherwise are specified.
Animals and study design
Female BALB/c mice (n = 90; 25–38 g) obtained from the University Centre of small animal breeding. The mice were placed in sterile cages under regulated conditions of temperature (22℃ ± 3℃), humidity (55% ± 5%) and 12 h/12 h day/night cycle. The mice were given free provided with adequate food and drinking water through the entire period of study. Study design and protocols were approved by the Ethical Committee of the Institute (approval number: TXY2015831). The animals were handled and housed as per the guidelines of National Animal Welfare Law of China and in compliance with the US National Institutes of Health Guidelines for the Care and Use of Laboratory Animals [28].
The mice were acclimatized to in-house conditions for 7 days prior induction of asthma. The mice were subjected to allergic asthma by OVA sensitization and inhalation as described Oh et al. [29] with slight modifications. In phosphate-buffered saline (PBS), 500 µg/ml of OVA was mixed with equal volumes of 10% (w/v) of aqueous alum. The pH was adjusted to 6.5 and the mixture was incubated for 1 h at room temperature. Following incubation, the mixture was centrifuged at 750 g for 5 min. The OVA/alum pellet was resuspended in distilled water to its original volume. On day 1, the animals were administered OVA at 100 µg (0.2 ml of 500 µg/ml OVA in normal saline) intraperitoneally (i.p.). The mice were then exposed to 250 µg OVA on day 8 (100 µl; 2.5 mg OVA/ml). On days 15, 18, and 21, the animals were challenged with OVA at 125 µg (50 µl of 2.5 mg/ml solution) intranasally as described by Oh et al. [29]. Mice were exposed to ketamine-xylazine anesthesia via i.p. (0.2 ml ketamine at 0.44 mg/ml and xylazine at 6.3 mg/ml in normal saline prior OVA challenge). Magnolol at 12.5, 25, 50 mg/kg body weight was administered to treatment group mice everyday via oral gavage, starting from 1st day of OVA exposure to day 21. On the days of OVA challenge magnolol was administered 60 min prior before each injection. Mice that received dexamethasone (Dex) at 2 mg/kg dose via i.p. an hour prior every OVA injection served as positive control. The control mice were not challenged with OVA and they received equal volumes of normal saline for magnolol.
Collection and analysis of bronchoalveolar lavage fluid (BALF)
Twenty-four hours following the last OVA challenge, the mice were sacrificed by pentobarbital overdose (50 mg/kg body weight) and tracheotomy was done. Ice-cold PBS (0.5 ml) was infused into a lung and BALF was collected via tracheal cannulation. BALF was collected by three successive aspirations (total volume 1.5 ml) [30] and then centrifuged (4℃, 250 g) for 5 min. The supernatant collected was stored (−70℃) until use. The supernatant was used for assessment of cytokines.
Cytokines IL-4, IL-5, IL-6, IL-13, IL-17A and IFN-γ in the BALF were determined by ELISA. The kits for analysis of IL-6, IL-17A and IFN-γ were purchased from R&D Systems (Minneapolis, MN, USA). ELISA kits for IL-4, IL-5 and IL-13 were purchased from Biolegend (San Diego, CA, USA). The analysis was performed as per manufacturer's protocol. Further, for determination of differential cell counts, cell pellets were re-suspended in PBS and stained with Wright–Giemsa stain (Nanjing Jiancheng Bioengineering Institute, Nanjing, China).
Determination of OVA-specific IgE in serum and BALF
The serum separated from whole blood by centrifugation (10 min; 3,000 revolutions per min; 4℃) was used for determination of OVA-specific IgE by ELSIA using LEGEND MAX™ Mouse OVA specific IgE kit (BioLegend) as per manufacturer's instructions.
Flow cytometric analysis of Th17cells
Th17 cell population was determined by flow cytometric analysis. Spleen excised from the mice of different experimental groups was placed in RPMI1640 media (Gibco, Grand Island, NY, USA). The spleen was disrupted using a cell disrupter (Beckman, Brea, CA, USA) to prepare a single cell suspension. The cells were washed in PBS. The cell pellet separated from BALF was resuspended in FBS and the supernatant was discarded. The cell pellet prepared from BALF and the cells of the spleen were incubated for 30 min with fluorescein isothiocyanate antihuman CD4 (eBioscience Co., San Diego, CA, USA) at 4℃. The cells were then fixed and permeabilized as per manufacturer's protocol, and were then stained with phycoerythrin anti-mice IL-17A (eBioscience Co). The cells were analyzed for Th17 cell population by flow cytometry (FACS, Calibur instrument with CellQuest software; BD Biosciences, Mountain View, CA, USA).
Determination of airway hyperresponsiveness
Functional changes of the airway following OVA challenge was assessed using methacholine. AHR to aerosolized methacholine was assessed within 24 h following final exposure to OVA. Buxco's modular and invasive system (Buxco Electronics Inc., Wilmington, NY, USA) was used to determine changes in lung dynamic complianc (Cdyn) and airway resistance (RI). Cdyn and RI in response to increasing concentrations of methacholine were recorded as described by Pichavant et al. [31]. In brief, the mice were tracheostomized and cannulated and laid in a whole-body plethysmograph chamber that was connected to the ventilator. The mice were administered methacholine at 3.125, 6.25, 12.5, or 25 mg/ml via a jet nebulizer into the head chamber. The values are expressed as percentage of the respective basal values as described by Glaab et al. [32].
Histologic analysis
Following collection of BALF, lungs were excised and fixed in 10% formalin, paraffinized and uniform sections (5 µm) were cut and stained with hematoxylin and eosin (H&E) to examine cellular infiltration into the peribronchial connective tissues. Cell count based on a five-point scoring system was done as described by Duan et al. [33]. The cell counts were scored as: 0, no cells; 1, a few cells; 2, a ring of cells 1 cell layer deep; 3, a ring of cells 2 to 4 cell layers deep; and 4, a ring of cells more than 4 cell layers deep.
Western blotting
The lung tissues were homogenized in liquid nitrogen and incubated in cell lysis buffer containing protease and phosphatase inhibitors (Roche, Basel, Switzerland) to obtain protein extracts. The total protein content present was determined using protein assay kit (Bio-Rad Laboratories, Hercules, CA, USA). Equal concentration of the protein samples (50 µg) were loaded on to 10% to 12% sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) gels and subjected to electrophoresis. The bands obtained were transferred onto a polyvinylidene difluoride membrane (Millipore, Temecula, USA). The membranes were then blocked with 5% non-fat dry milk and incubated with primary antibodies overnight at 4℃. The membranes were further incubated with peroxidase-conjugated secondary antibodies at room temperature for 1 h. The immunoreactive bands were visualized and analysed using a chemiluminescence system (Amersham Bioscience, Buchinghamshire, UK). The band intensities were normalized to β-actin expression that was used as internal control.
RESULTS
Magnolol reduces airway hyperresponsiveness
Asthma is characterized by non-specific hyperresponsiveness of the airway in response to various allergens and contractile agonists as OVA. In the study, effects of magnolol (12.5, 25, and 50 mg) on AHR was determined by measuring Cdyn and RI in response to increasing doses of methacholine in mechanically ventilated mice. RI is defined as the pressure driving respiration by air flow. Lung compliance is the distensibility of the lungs and is defined as the change in lung volume due to change in lung pressure. Here, OVA sensitization and challenge induced AHR, as reflected by significantly (p < 0.05) high RI and low Cdyn (Fig. 1) v.s. control mice not exposed to OVA. Interestingly, magnolol administration at the tested doses significantly (p < 0.05) reduced RI and restored Cdyn in response to methacholine. Also, magnolol at 50 mg exhibited maximum protective effects as against lower doses more positive results and the effects were close to that of positive control, Dex.
Magnolol inhibited inflammatory cell infiltration into BALF
Inflammatory responses are well established in allergic conditions as asthma. The effects of magnolol on cellular infiltration into the BALF were determined. OVA-sensitized mice, exhibited severe infiltration of eosinophils and leukocytes in the BALF (Fig. 2). Magnolol supplementation significantly (p < 0.05) inhibited cellular infiltration following OVA sensitization and challenge as reflected by the decrease in number of eosinophils, macrophages and neutrophils in the BALF. Further 50-mg magnolol exhibited suppressive effects in line with standard drug, Dex.
Magnolol reduced levels of OVA specific IgE
Raised expressions of allergen-specific IgE on the surface of mast cells in response to an allergen challenge is regarded as one of the early events in asthma. OVA-specific IgE levels in the serum and BALF was determined 24 h following the last OVA challenge. OVA exposure caused a significant increase (p < 0.05) in the serum and BALF levels of OVA-specific IgE v.s. control mice. Interestingly, magnolol treatment to OVA-sensitized animals significantly (p < 0.05) reduced the levels of OVA-specific IgE in both serum and in BALF (Fig. 3). Magnolol of 50 mg reduced the levels to almost 5 times in serum and 4 times in BALF.
Magnolol restores Th1/Th2 cytokine balance and regulates Th17 cytokine levels
Th1/Th2/Th17 cytokines play vital roles in allergic asthma. The effects of magnolol the cytokine levels in the BALF were determined. Th2 cytokines (IL-4, IL-5, and IL-13) and Th17 cytokines-IL-17A and IL-6 were observed to be significantly raised (p < 0.05) following OVA sensitization and challenge (Fig. 4A, B). However, IFN-γ, the Th1 cytokine (p < 0.05) was markedly decreased. Magnolol caused a significant (p < 0.05) decrease in the levels of Th2 and Th17 cytokines along with a noticeable increase in IFN-γ in a dose-dependent manner. A 50-mg dose of magnolol decreased IL-17A and IL-6 from 104 and 165 pg/ml to 41 and 48 pg/ml, respectively; while IFN-γ levels increased from 312 to 519 pg/ml. The observations indicate the effective restoring of the Th1/Th2 balance and Th17 cytokine levels in line with the effects of Dex.
The effect of magnolol on Th17 cell population
Th17 cells are known to be involved in inflammatory responses following allergen exposure. The relative proportion of Th17 cells was determined in the spleen of mice following OVA challenge. Flow cytometry showed dramatically (p < 0.05) increased number of Th17 cells v.s. normal control (Fig. 5). The Th17 population was found to be raised 6.5 folds as compared to control. Administration of magnolol at all the 3 tested doses significantly (p < 0.05) decreased Th17 cells. The Th17 cell population was reduced to near normal counts with 50-mg magnolol. This reduction in Th17 cell population by magnolol could be responsible to the decreased levels of cytokines IL-6 and IL-17A.
Magnolol restores lung architecture following OVA challenge
Histological analysis of the lung tissues following OVA challenge was determined by H&E staining. The lung tissues revealed marked infiltration of inflammatory cells into the peribronchiolar and perivascular connective tissues upon OVA-sensitization and challenge (Fig. 6). Magnolol administration was observed to markedly inhibit the infiltration inflammatory cells-eosinophils and neutrophils into the lung tissues. Eosinophilia observed on OVA challenge was substantially decreased in line with cellular infiltration into BALF. Also, magnolol considerably restored lung architecture to near normal with 50 mg dose exerting maximum protective effects. Mice treated with 50 mg magnolol revealed nearly normal lung histology.
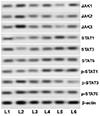
Activation of JAK-STAT pathway following OVA-sensitization and challenge
JAK-STAT pathway is well documented in the pathogenesis of asthma. The pathway is known to affect the Th1/Th2 differentiation and imbalance [12]. Here the effect of magnolol on the major proteins of the pathway was evaluated by western blotting. OVA challenge markedly raised the expressions of JAK1, JAK2, JAK3 and p-STAT1, p-STAT3, p-STAT6 suggesting activation of the pathway (Fig. 7). The expressions of total STAT1, STAT3, and STAT6 were also increased noticeably. However, magnolol treatment significantly down-regulated the phosphorylated levels of STAT1, STAT3, and STAT6 indicating down-regulation of the pathway. Further, the expressions of JAK1, JAK2, and JAK3 were noticed to be considerably reduced. Magnolol-mediated reduced levels of ILs could have in part aided in down-regulation of the JAK/STAT pathway.
Magnolol modulated Notch signaling
Notch signaling is well documented in the differentiation and activation of T cells. Here the OVA challenged mice presented with significantly (p < 0.05) up-regulated expressions of Notch 1 receptor along with the ligands-Jagged1 and Jagged2 (Fig. 8). We observed significant (p < 0.05) down-regulation in Notch signaling in magnolol administered mice. The expression of Notch 1 was brought to almost normal levels following administration of 50-mg magnolol. The observations reveal that effective down-regulation of JAK/STAT/Notch signaling by magnolol that is suggestive of the potential anti-inflammatory effects (Fig. 9).
DISCUSSION
Asthma is a common allergic disease affecting about 7% to 13% of the global population [34]. The pulmonary inflammatory response to allergens in asthma causes AHR, hypersecretion of mucus, bronchoconstriction and airway remodeling leading to shortness of breath, wheezing, and chest tightness [3536]. It is a huge burden that affects the quality of living of asthma patients. Corticosteroids are currently used main therapeutic drugs for asthma [3738]. Nevertheless, these drugs do not completely cure or modify the progression of asthma. The symptoms return once the treatment is terminated and further, continued use of these drugs may often lead to systemic and as well local side effects [3940]. Thus, identification of novel effective therapeutics is of immense clinical value. In recent years the use of plant-derived compounds as alternative medicine in the treatment of various clinical conditions is increasing [414243]. In this study, we assessed the effects of magnolol, in OVA-induced rodent model of experimental asthma.
AHR is one of the hallmark characteristics in asthma presenting with bronchoconstriction due to inflammatory responses and hypertrophy of airway smooth muscles eventually resulting in impaired function of the lungs [44]. Airway functioning following OVA challenge was assessed by measuring airway RI and lung compliance. OVA exposure was found to cause increase airway RI and decreased compliance of the lungs. Magnolol improved lung function as revealed by restored lung compliance and decreasing airway RI in response to varied concentrations of methacholine.
The early responses to allergens in asthma are known to be IgE dependent. Higher levels of IgE has been found to be responsible for degranulation of mast cells and bronchial constriction [45]. Further, allergic airway responses in the late phase of asthma is associated with the infiltration of inflammatory cells particularly eosinophils and lymphocytes into the bronchial tissues [4647]. Eosinophilia is a characteristic feature in asthma. The inflammatory cells produce cytokines and chemokines that induce AHR [48]. Magnolol was found to effectively attenuate the migration of inflammatory cells into the lung tissues. Magnolol strikingly reduced eosinophilia and inflammation score and as well as OVA-specific IgE levels in serum and BALF. Histological examination also revealed significantly restored lung architecture following magnolol treatment.
The crucial role of T lymphocytes in asthma pathogenesis is well documented, through the release of cytokines [49]. The Th2 cytokines—IL-4, IL-5, and IL-13 mediate several hallmark features including airway inflammation and AHR. Up-regulated levels of Th2 cytokines are observed in asthma [505152]. The raised levels contribute to inflammatory cell infiltration, eosinophil activation, raised IgE production and mucus hypersecretion [5354]. Cytokine, IL-4 is a key factor that is involved in isotype switching to IgE in B cells [55]. IL-5 is pivotal for eosinophil activation and survival [6] while, IL-13, is involved in eosinophil infiltration into lung tissue. IgE levels are known to dependent on IL-4 and thus the levels considered as an additional index of Th2 cytokines in asthma. Thus, IgE could be considered as a key target in developing novel anti-asthma drugs. Reducing IgE levels could be effective in asthma therapy.
Th17 cells are important subset of T-helper cells that exerts crucial roles in the pathogenesis of inflammatory diseases such as asthma [910]. Studies with rodent models of asthma have reported that Th17 cell-derived cytokines as- IL-17A and IL-17F, are implicated in recruitment and activation of antigen-induced neutrophils. Th17 cytokines were also found to enhance Th2-mediated recruitment of eosinophils into the bronchial tissues [565758]. Raised levels of Th17 cell-derived cytokines were noticed in the lung tissues and in BALF of patients with asthma and further, the levels of Th17 cytokines were reported to correlate positively with severity of AHR [5960]. Thus, the increased levels of the IL-17A as observed following OVA challenge indicates raised cellular infiltration and inflammation. Reduced expression of Th2 and Th17 cytokines and as well IgE levels by magnolol reveals the efficacy of magnolol in inhibiting inflammatory responses. Previous studies with plant-derived compounds as curcumin exerted anti-asthmatic effects via significantly reducing cytokine levels in OVA-induced asthma [61].
Also, it is well documented that Th1 cell derived IFN-γ suppresses Th2 cell-mediated immune responses. IFN-γ is involved in IgG2α class switching. Magnolol treatment markedly raised IFNγ levels [8]. The raised IFNγ levels illustrates enhanced Th1 responses that in part could have contributed to decreased IgE levels.
The JAK-STAT signaling pathway is well documented in Th2 differentiation predominantly and in Th1/Th2 balance [12]. JAKs belong to the family of tyrosine kinases and are regulators of signaling through cytokine receptors. Out of the 4 JAKs (JAK1, JAK2, JAK3, and TYK2) among which JAK1 and JAK3 are crucial in the initiation of inflammation [62]. Ashino et al. [12] reported that a selective inhibitor of JAK1/3 inhibited Th2 cell differentiation, down-regulated phosphorylation of STAT5 and 6, prevented AHR, mucus hypersecretion, and production of Th2 cytokine. Upon activation by phosphorylation, the active STATs translocate into the nucleus where they act as transcription factors to regulate gene expression [636465]. STAT3 that signals downstream of IL-6 has been found to be critical for the differentiation of Th17 cells [6667]. STAT6 regulates several features of lung inflammatory responses including infiltration of inflammatory cells in to the airway, Th2 cell differentiation, and IgE production [68]. STAT1 is associated with IL6 and IL11 [14]. The observed elevated expressions of phosphorylated forms of JAK1, JA2, JAK3, STAT1, STAT3, and STAT6 reflect activation of the JAK/STAT pathway. Magnolol-mediated decrease in phosphorylated levels of JAKs and STAT1, STAT3, and STAT6 illustrates down-regulation of the pathway as a mechanism against asthma.
Notch signaling has also been reported to participate in the activation and proliferation of T cells [697071]. In line with earlier studies raised expressions of Notch1 receptor and Jagged1 and Jagged2 ligands following OVA challenge [4272] were observed. Guo et al. [21] reported that blocking of Notch1 by Notch1-specific small interfering RNA, the levels of IL-4 decreased while IFN-γ is increased, suggesting role of Notch1 signal in Th1/Th2 differentiation in asthma [21]. Magnolol effectively down-regulated the expressions of Notch1 receptor and Jagged1 and Jagged2. These observations suggest the efficacy of magnolol in regulating T cell differentiation and activation, thus aiding in decreased cytokine levels and inflammation.
The results indicate the efficacy of magnolol in reducing airway inflammation and AHR via JAK-STAT and Notch signaling and by regulating Th1/Th2/Th17cytokine levels. Thus, suggesting magnolol, as a potent candidate drug in asthma therapy. However, the compound has to be explored further to understand the possible mechanisms involved.







 XML Download
XML Download