

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Resistance training (RT) precipitates an anabolic environment by inducing both hormonal and molecular variations in skeletal muscle. RT contribute to enhancement of insulin, insulin-like growth factor-1 (IGF-1), and myokines, which are well known as anabolic hormones [1,2,3]. Specially, RT-induced elevation in IGF-1, which can regulate muscle hypertrophy and muscle strength, has been strongly associated with myokine levels [4]. Numerous studies indicate that the novel myokine irisin, which is a cleaved and secreted form of fibronectin III domain-containing 5 found in organs including brain, heart, adipose tissue, and skeletal muscle, provokes browning of white adipose tissue a process controlled by uncoupling protein 1 and peroxisome proliferator-activated receptor-gamma co-activator-1 alpha [5]. Some authors have suggested that exercise-mediated increase in circulating irisin was strongly related with improvement of body fat mass, insulin resistance, and IGF-1 [6,7,8,9,10]. Furthermore, a study by Bostrom et al. [11] suggested that the plasma level of circulating irisin increased after 10 weeks of endurance training. In contrast, Norhein et al. [12] and Moraes et al. [13] found a decrease in the plasma concentration of irisin after 12 weeks of training and 6 months of resistance exercise, respectively. Therefore, the precise role of irisin in exercise-related physiological change is currently controversial [14].
The benefits of nutritional strategies to increase muscle strength have been widely investigated for potential improvements to health outcomes in humans [15,16,17,18,19]. In this context, supplementation with phytochemical compounds has received significant attention for its potential applications to resistance training. Ursolic acid (UA) is an isomer of oleanolic acid and a lipophilic triterpenoid compound primarily found in apples and many herbs [20]. Recently, UA has been demonstrated to have important biological roles including anti-fat accumulation [21], anti-insulin resistance via IGF-1 [6], anti-muscle atrophy [22], anti-cancer [23], anti-oxidation [24], and anti-inflammatory effects [25]. Above all, the underlying mechanism of UA is the enhancement or maintenance of skeletal muscle mass and function in vivo and vitro [22,25,26,27]. However, it is not clear whether increase in skeletal muscle mass is associated with signaling via the same IGF-1-irisin-pathway found to promote skeletal muscle strength in existing clinical studies. To date, several studies have assessed the impact of UA supplementation on muscle atrophy or obesity, but no reports have elucidated the biological effect of UA supplementation on human subjects involved in RT. For that purpose, we determined differences in body composition, fasting glucose, insulin, IGF-1, irisin levels, and muscle strength following RT with or without concomitant UA supplementation. We hypothesized that UA-induced elevation of serum irisin would promote body fat loss and improvements in skeletal muscle development during RT in men.
Go to : 
METHODS
Study design
The participants of this study performed an exercise program that included supervised RT with or without UA supplementation for 8 weeks. Body composition, blood parameters and muscular strength were determined at baseline and after this program. All participants signed written informed consent approved by the Pusan National University Institutional Research Board (PNU IRB/2014_42_HR).
Subjects
Twenty-four Korean men with over 3 years of RT experience were recruited from Dong Myoung University by newspapers and e-mail for this study. All respondents completed a family history and detailed health history questionnaires. Respondents were excluded from this study (n=8), if they met exclusion criteria including diagnosis of chronic disease including cardiovascular disease, hypertension, cancer, kidney dysfunction, or musculoskeletal dysfunction during the previous 3 months. Sixteen healthy participants (age: 29.37±5.14 years) were ultimately selected to participate in this study. All eligible participants were nonsmokers and had not taken medications or steroid therapy within 1 year prior to or during this study period. Participants were randomly divided into the following interventions: RT with placebo supplementation (RT; age: 28.71±5.76 years, n=7) and RT with UA supplementation (RT+UA; age: 29.88±4.91 years, n=9). Before the investigation, participants stopped consumption of other ergogenic aids immediately and were subjected to a 5-week washout period.
Resistance training
Participants performed the RT program for 8 weeks using free weights and machines under the supervision of a trained instructor, who directed stretching exercises as a warm up to prevent soft tissue injuries and ensure subject safety. The accuracy of a prediction equation to estimate 1 repetition maximum intensity (1RM) until executed to fatigue was evaluated at each exercise session before starting the program [27]. This RT program consisted of a total of 26 exercise types, with 13 upper-body (chin-up, lateral pull-down, bent over/up-right row, bench press, fly, cable cross-over, shoulder press, lateral raise, biceps curl, concentration curl, barbell elbow extension, and kick back) and 13 lower-body training exercises (squat, leg extension, leg press, sissy squat, leg curl, dead lift, stiff leg dead lift, lunges, back extension, seated/standing calf raise, sit-up, and abdominal crunch) to strengthen the extremities. Participants performed 5 sets of 10~15 repetitions at 60~80% of the 1RM with a 60~90-s inter-set rest, 6 times/week for 8 weeks.
Ursolic acid supplementation
The placebo group was provided with 3 capsules daily containing 450 mg of guar gum, and the UA group received 3 capsules daily containing 450 mg of UA from rosemary leaf extract (Labrada, Houston, Texas, USA). The capsules had the same color, shape, and size, and participants took 1 capsule (150 mg) after each of 3 meals. We monitored the participants by questionnaire via e-mail, cell phone, and visits to the laboratory to confirm compliance with supplementation throughout the study.
Anthropometric and biochemical parameters
Blood samples were collected before and after the 8-week intervention. Body weight and height were measured with the participants wearing light clothes and no shoes. Body composition including body weight, body fat percentage and lean body mass was measured before and after the 8-week intervention using a multi-frequency electrical impedance analyzer (X-scan Plus II, Jawon Medical, Seoul, Korea). Body mass index was calculated as body weight (kg) divided by the square of height (m). Blood samples were collected from the antecubital vein of the arm into vacutainer tubes (BD Bioscience, San Jose, CA, USA) before and after the 8-week intervention. Blood samples were separated by centrifugation at 1,500×g and 4℃ for 15 min and stored frozen at -80℃ until assayed. Fasting serum glucose level was measured using an automated glucose analyzer (ADVIA 1650, Bayer). Serum insulin (Mercodia, Winston Salem, NC, USA) and irisin (Phoenix Pharmaceuticals, Inc., Burlingame, CA, USA) were assessed using enzyme-linked immunosorbent assay kits. IGF-1 was determined by chemiluminescence immunoassay using the IGF-1 assay kit (Siemens Corporation, UK).
Isokinetic muscle strength (peak torque)
Participants' maximal muscle strength was evaluated at baseline and after 8-week intervention during a laboratory visit. They were seated on a Human/Norm Testing and Rehabilitation System (Computer Sports Medicine, Inc., MA, USA, 2006). Their chests, hips, and thighs were fixed using a belt to minimize the influence of other muscles. The axis of rotation of the knee joint was aligned with that of a dynamometer. After the dynamometer was fixed to the ankle with a strap, anatomical zero was set, and the movable range was set with extension at 0° and flexion at 110°. The participants were provided with sufficient explanation of the test procedure and asked to repeat the flexion/extension exercise twice to adapt to the test before the measurement. Peak torque was determined by completion of 1 set of 3 flexion/extension 60°/s. Participants had continuous verbal encouragement from supervisors during all tests, and as measurement criteria we selected peak and average torque at each repetition.
Statistical analysis
For all variables, data were expressed as mean±SD and analyzed using PASW Statistics for Windows software version 19.0 (SPSS, version 19.0 for Windows; SPSS, Inc., Chicago, Illinois, USA). The effects of RT and UA supplementation on body composition, blood parameters and muscle strength were tested using two-way ANOVA with repeated measures (2 [groups]×2 [time points]). When a significant group-by-time interaction was detected, within-group differences between mean values were analyzed using a paired t-test. The level of significance was considered p<0.05.
Go to :
RESULTS
The characteristics of the study participants are shown in Table 1. At baseline, no significant differences in age, height, weight, body mass index, body percentage, or lean body mass were observed between the RT and RT+UA groups. After the RT+UA intervention, body fat percentage was significantly decreased compared to that at baseline in the RT group (p<0.001). Although lean body mass demonstrated a slight increase compared with baseline levels (p>0.05), no significant changes were observed in any characteristics of body composition.
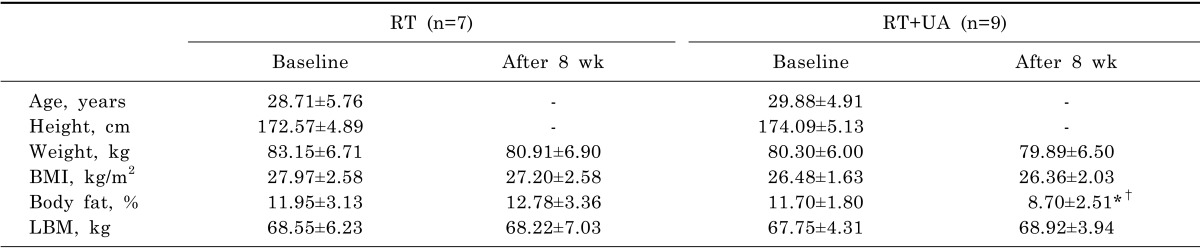
Blood parameters of the study participants are shown in Table 2. At baseline, no significant differences in blood parameters were observed between the RT and RT+UA groups. After RT+UA intervention, IGF-1 and irisin levels were significantly increased compared to baseline in the RT group (p<0.05), while glucose and insulin did not change in either group.
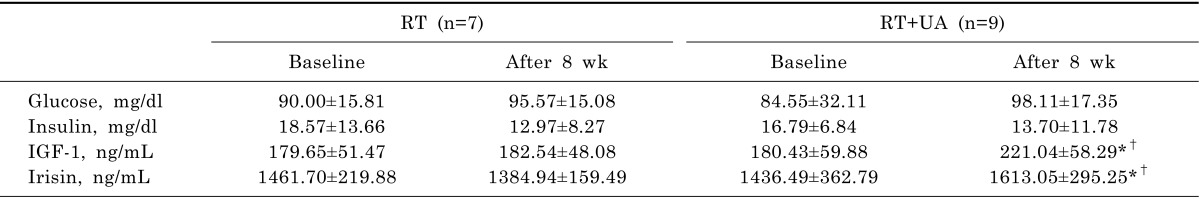
Isokinetic maximal muscle strength of the study participants is shown in Table 3. At baseline, no significant differences in muscle strength were observed between the groups. After RT+UA intervention, maximal right extension (p<0.01), maximal left extension (p<0.01), maximal right flexion (p<0.05), and maximal left flexion (p<0.001) were significantly increased compared to baseline. There were significant group-by-time interactions for maximal right extension (p<0.01), maximal left extension, maximal right flexion, and maximal left flexion (p<0.05) indicating that the increases in isokinetic maximal strength in the RT+UA groups significantly differed from the non-significant changes from baseline in the RT group.
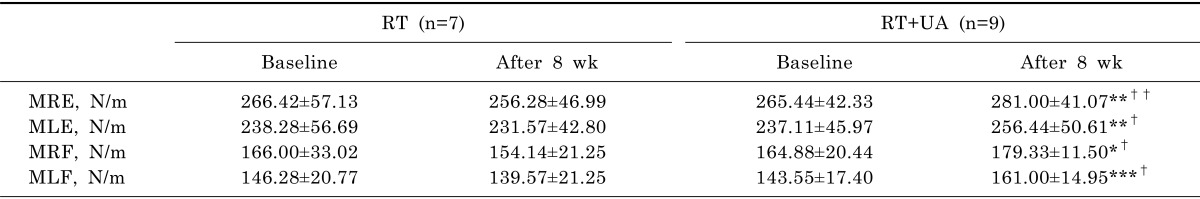
Table 3
Maximal muscle strength of participants before and after study interventions
Values are expressed as mean±standard deviation. MRE, maximal right extension; MLE, maximal left extension; MRF, maximal right flexion; MLF, maximal left flexion. Within-group differences were analyzed by paired t-test (*p<0.05, **p<0.01, ***p<0.001). Analysis of variance was used for group-by-time interactions (†p<0.05, ††p<0.01).
![]()
Go to :
DISCUSSION
In the present study, we demonstrate for the first time that prolonged RT with UA supplementation in men strikingly enhanced physiological changes including a decline in body fat percentage, increase in IGF-1 and irisin levels, and increase in muscle strength. In contrast to, no change in skeletal muscle mass was observed. These findings indicate that RT with UA supplementation may represent a better strategy for increasing skeletal muscle strength via an improvement in plasma IGF-1 and irisin compared to RT alone. Therefore, it appears that UA supplementation can exert a beneficial effect on a combined exercise program, although it may have a mismatched effect on skeletal muscle mass.
Relevant strategies for improving exercise performance include not only positive training methods [28,29,30], e.g. resistance exercise, but also nutritional protocols [15,31]. Many edible fruits and herbs contain UA, which is a bioactive phyto-constituent [20,32,33,34,35] and is also used in anti-obesity herbal medicines [21]. UA appear to strongly inhibit body fat gain in obese humans and animals [26,27]. Similar to these previous results, we observed significant changes in body fat percentage, but no changes in body weight (Table 1). These results suggest that UA supplementation lowered the body fat percentage in men during the study intervention, whereas RT alone did not affect body fat percentage. Therefore, the significant decrease in body fat percentage without change in body weight induced by UA supplementation may be explained by favorable changes in body composition including a decrease in body fat mass [36].
A recent study suggested UA supplementation as a new therapeutic application for skeletal muscle hypertrophy through activation of IGF-1 signaling [6]. In addition, Ogasawara et al. [26] found that UA supplementation stimulated skeletal muscle hypertrophy after RT through Akt-independent activation of mammalian target of rapamycin complex 1 activity, which is known as an important regulator of muscle protein synthesis and hypertrophy [37]. For these reasons, we hypothesized that UA supplementation may improve skeletal muscle hypertrophy via IGF-1 signaling during RT, however, we did not observe substantial gains in lean body mass (Table 1), although a slight, non-significant upward trend was noted. A previous study reported that an increase in lean body mass can stimulated hormones or myokines, which are associated with increased skeletal muscle mass [38]. Additionally, IGF-1 is strongly associated with skeletal muscle mass; is an important hormone involved in skeletal muscle hypertrophy [39]; and regulates cell proliferation, differentiation [40], and satellite cell function during muscle regeneration [41]. To support our hypothesis that UA exerts effects through increased IGF-1, we observed a significant increase in IGF-1 in the RT+UA group compared to the RT group (Table 2). It appears that UA supplementation may play a role in augmenting IGF-1 without a change in skeletal muscle hypertrophy. Many studies have reported that RT alone [42,43] and UA supplementation [22] significantly impact stimulated IGF-1. These findings indicate that an enhancement of the increase in IGF-1 could be expected in skeletal muscle hypertrophy. However, our training program was not associated with marked skeletal muscle hypertrophy. This discrepancy may be due to our study population, since the men already demonstrated muscle hypertrophy, and thus a further significant increase in muscle mass following 8 weeks of RT alone and RT+UA is not easily achieved. In addition, the adaptation of skeletal muscle hypertrophy, which can lead to the stimulation of other phenomena, may depend on exercise training intensity, frequency, and the duration of physical activity [44]. Therefore, further investigation is needed to consider exercise training protocols and verify the mechanisms underlying UA-associated skeletal muscle hypertrophy in resistance trained-men.
Skeletal muscle has been recently identified as an endocrine organ that can regulate several physiological and metabolic pathways in response to muscle contraction [45]. Several myokines [46] and IGF-1 may be involved in inducing skeletal muscle hypertrophy or suppressing skeletal muscle atrophy [47]. One of these myokines is irisin, which plays a role in energy generation in the skeletal muscle [48]. The exercise-mediated increase in circulating irisin was strongly related with improvement of IGF-1 [6,7,8,9,10]. To clarify the inconsistency in previous findings on irisin, we measured the plasma level of irisin during RT+UA supplementation and found a significant increase in irisin level in the RT+UA group compared to the RT group (Table 2). This finding is the first reported evidence that UA administration leads to increased irisin and that RT alone does not affect this myokine in men. This result raises the possibility that blood levels of IGF-1 and irisin are regulated differently, but the mechanisms underlying their regulation remain unknown.
Finally, we hypothesized that the increase in IGF-1 and irisin would improve maximal muscle strength, since skeletal muscle strength is a quality indicator of skeletal muscle function. UA supplementation during RT significantly enhanced isokinetic muscle strength (Table 3). This result provides another indication that UA may play an important role in muscle strength as well as muscle mass. The finding highlights a novel possibility for obtaining more muscle strength through UA supplementation in men that may perhaps be applicable to other populations who practice RT, such as individuals with muscle atrophy due to aging and/or disease. It is difficult to directly connect increased muscle strength with irisin level following UA supplementation and we cannot exclude the possibility that the increased muscle strength may be related to irisin in combination with IGF-1 or IGF-1 alone. However, these findings suggest that irisin may be involved in exercise-mediated skeletal muscle contraction by increasing the ability to generate maximal muscle force (i.e. muscle strength). The increase in maximal muscle strength in the RT+UA group is highly relevant to health-related fitness benefits, even in well-functioning subjects, since muscle strength helps to maintain a high level of physical capacity. To our knowledge, no other study has reported independent muscle size and muscle strength responses to UA supplementation in resistance-trained men during RT. The relevance of specific mechanisms underlying these effects of myokines requires further study.
The present study has some limitations. Although significant results were observed in the relatively small number of participants, significant correlations could not be detected in this study. Future studies should aim for a larger number of participants. We provided education about diet to all participants to avoid other diet effects during the study; however, we could not confirm detailed dietary patterns during the study period. Finally, our study was performed in healthy men. In future investigations, the biological effect of UA intervention alone or combined with RT should be performed in individuals with disease states such as muscle weakness and age-and/or disease-related muscle atrophy to further elucidate the muscle strength-enhancing properties of UA.
In conclusion, our results show that an 8-week RT program with UA supplementation can decrease body fat percentage and increase IGF-1, irisin, and skeletal muscle strength without affecting muscle mass in men. Thus, this approach appears to be a promising strategy to improve skeletal muscle strength in men. However, the underlying molecular mechanisms involved in the increase of IGF-1, irisin, and muscle strength achieved through UA supplementation require further investigation.
Go to :
 XML Download
XML Download