

 PDF
PDF ePub
ePub Citation
Citation Print
Print


ABBREVIATIONS
AAS
α-asarone
CORT
corticosterone
EMP
elevated plus maze
HBT
hole-board test
CRF
corticotrophin-releasing factor
TH
tyrosine hydroxylase
LC
locus coeruleus
BDNF
brainderived neurotrophic factor
CNS
central nervous system
GAD
generalized anxiety
PD
panic disorder
SAD
social anxiety disorder
PTSD
post-traumatic stress disorder
HPA
hypothalamicpituitary-adrenal
NO
nitric oxide
SD
Sprague-Dawley
DZP
diazepam
ELISA
enzyme-linked immunoassay
ABC
avidin-biotinperoxidase complex
DAB
3,3'-diaminobenzidine
RT-PCR
reverse transcription-polymerase chain reaction
GAPDH
glyceraldehydes-3-phosphate dehydrogenase
ANOVA
analyzed by analysis of variance
INTRODUCTION
Anxiety, an aversive emotional state, is the most common mental disease of the central nervous system (CNS), more serious disorder to the health, and it contributes to an ever increasing healthcare burden worldwide. It consists of a powerful emotional component associated with fearful thoughts and a physiological response [1]. Some studies have reported that anxiety is the most frequent psychiatric disorder, closely followed by mood disorders [2]. Despite a steady increase in the development of anxiolytics for anxiety, the prevalence of this disorder remains stable, which may be attributed to our poor neurobiological understanding of its pathophysiology or to the inconsistent efficacy of current treatments [1]. Anxiety disorders include generalized anxiety (GAD), panic disorder (PD), social anxiety disorder (SAD), and post-traumatic stress disorder (PTSD) [3]. They result from exposure to stressful life events that evoke fear, helplessness, and horror. Indeed, chronic exposure to stressful life events is an established and important risk factor for the development and maintenance of many psychological or mental disorders, including anxiety, in humans [4]. The behavioral and physical responses to stress are especially initiated by activation of the hypothalamic-pituitary-adrenal (HPA) axis, and this results in the release of catecholamines and stress hormones such as corticosterone (CORT) [5].
Many studies have shown that stimulation and sustained action of the HPA axis are closely associated with the development of stress-related psychosomatic disorders, which produce serious changes in affective behavior that are indicative of or consistent with anxiety-like symptoms [6,7]. Repeated CORT administration may lead to profound maladaptive changes in anxiety-like behavior, thereby mimicking human mental disorders such as anxiety [8]. Previous studies have shown that chronic exposure to CORT increased the plasma CORT concentration, inhibited exploratory behavior, enhanced freezing behavior in fear conditioning, and induced changes in c-Fos expression in the hypothalamus, amygdala, and hippocampus [9]. Thus, chronic CORT-induced physiological stress resulting from HPA axis dysregulation increases anxiety-like behavior [10].
Moreover, diazepam is generally prescribed medication for the treatment of several forms of anxiety, but this agent has prominent side effects, such as sedation, dependence, and cognitive and psychomotor impairment [3]. Thus, much attention has been given to the use of naturally occurring compounds and their formulations as alternative therapeutic agents for the treatment of psychiatric disorders including anxiety [11,12].
α-asarone (AAS, trans-1-propenyl-2,4,5-trimethoxy-benzene) is a major compound isolated from Acorus gramineus [13]. The compound has antioxidant, anticoagulant, antitumor, antiviral, and anti-inflammatory activities, suggesting potential value for medicinal use [14]. Indeed, there are many AAS products, including tablets, capsules, and injections, available on the market. Recently, AAS has been reported to improve multiple physiological actions, produce a variety of pharmacological actions in the CNS, and modulate immune system function [15]. However, some of the studies on the anticonvulsant activity of AAS yielded only marginal effects [13,16]. Several studies have shown that the administration of AAS produced weak anticonvulsant activity during maximal electroshock seizures and pentylenetetrazole-induced seizures [13], with antioxidant properties, such as those of superoxide dismutase, catalase, and glutathione peroxidase, against noise stress-induced changes in the rat brain [17]. In addition, AAS protected neurons against amyloid-beta (25-35)-induced neurotoxicity and spatial memory impairment by inhibiting the effects of nitric oxide (NO) overproduction in the hippocampus and temporal cortex [18]. Thus, AAS may act to alleviate anxiety-behavior in stress-related animal models. However, it remains unknown to date whether the therapeutic efficacy of AAS in alleviating anxiety-behavior following chronic stress-induced anxiety-like symptoms induced by repeated CORT injections in rats is mediated by the regulation of BDNF and TrkB. These observations resulted in the hypothesis that AAS may also be effective in improving stress-associated psychiatric conditions, such as anxiety.
Based on these observations, we sought to evaluate in the present study the efficacy of AAS administration for alleviating anxiety-related symptoms in rats exposed repeatedly to exogenous CORT. Anxiety-like behaviors were assessed by the elevated plus maze (EPM) test and the hole-board test (HBT). Additionally, the underlying neurobiological mechanism of these behaviors was investigated through evaluation of BDNF signaling regulation via TrkB expression in the brains of rats following repeated CORT injections.
METHODS
Animals
Adult male Sprague-Dawley (SD) rats weighing 200-220 g (6 weeks-old) were obtained from Samtako Animal Co. (Seoul, Korea). The rats were housed in a limited access rodent facility with up to five rats per polycarbonate cage. The room controls were set to maintain the temperature at 22±2℃ and the relative humidity at 55±15%. Cages were lit by artificial light for 12 h each day. Sterilized drinking water and standard chow diet were supplied ad libitum to each cage during the experiments. The animal experiments were conducted in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publications No. 80-23), revised in 1996, and were approved by the Kyung Hee University Institutional Animal Care and Use Committee. All animal experiments began at least 7 days after the animals arrived. The effects were made to minimize the number and suffering of animals.
Experimental groups
This study was designed to explore the efficacy of AAS administration for healing repeated CORT-induced anxiety-like behaviors in an animal model using behavioral and neurobiological methodologies. Different groups of rats, 5 or 6 animals per group, were used for drugs treatment and tests. All the experimental animals including control and drug-treatment groups were administration. The aim of food withdrawn prior to drug administration was just to ensure the bioavailability of drug. In other time periods, all animals had free access to food. The procedure and dose of CORT administration was performed as described in Mao et al. [19]. CORT (40 mg/kg, Sigma-Aldrich Chemical Co. St. Louise, MO, USA), which was dissolved in absolute ethanol and subsequently diluted in water to the final concentration of 10% ethanol, was administrated by subcutaneously (s.c) in a volume of 1 ml/kg once daily for 21 days [8,20]. This CORT dose was selected because it induces serum levels of the steroid comparable to those elicited by substantial stress [21]. The CORT and vehicle injections were given in the morning between 9 and 10 am once daily for 21 consecutive days. The standard doses of AAS in the rat and considering the long-term treatment used in the present study was based on previous study [13]. AAS (50, 100, 200 mg/kg body weight, Sigma-Aldrich Chemical Co.) and the positive drug diazepam (1 mg/kg, DZP; Roche, Welwyn Garden City, UK) were administered by intraperitoneally (i.p.) in a volume of 1 ml/kg 30 min prior to the CORT injection for 21 days. All drugs were freshly prepared right before every experiment. The following parameters were measured to monitor the effects of the development of psychosomatic disorders by exogenous CORT administration: changes of body weight gains (at the beginning step of exogenous CORT administration), and serum CORT levels (after repeated CORT-induced anxiety-like symptoms). Behavioral testing for anxiety-like behaviors were done rightly after the end of the chronic physiological stress protocol. All rats sequentially performed to take the EPM test and HBT on the 21th day after repeated CORT injection. After the behavioral testing and body weighting, rats were sacrificed and brain tissues were immediately collected for experiments or stored at -70℃ for later use.
Elevated plus maze test (EPM)
The EPM test is a widely used behavioral test to assess anxiogenic or anxiolytic effects of pharmacological agents. Animals conduct anxiety-like behaviors usually show the reductions both in the number of entries and in the time spent in the open arms, along with an increase in the amount of time spent in the closed arms in the EPM. The elevated plus test was conducted. This apparatus consisted of two open arms (50×10 cm each), two closed arms (50×10×20 cm each) and a central platform (10×10 cm), arranged in a way such that the two arms of each type were opposite to each other. The maze was made from black Plexiglas and elevated 50 cm above the floor. Exploration of the open arms was encouraged by testing under indirect dim light (2×60 W). At the beginning of each trial, animals were placed at the centre of the maze, facing a closed arm. During a 5-min test period, the following parameters were recorded: 1) number of open arm entries, b) number of closed arm entries, c) time spend in open arms, and d) time spent in closed arms. Entry by an animal into an arm was defined as the condition in which the animal has placed its four paws in that arm. The maze was cleaned with alcohol after each rat had been tested. The behavior in the maze was recorded using a video camera mounted on the ceiling above the center of the maze and relayed to the S-MART program (PanLab, Barcelona, Spain). Anxiety reduction, indicated by open arm exploration in the EPM, was defined as an increase in the numbers of entries into the open arms relative to total entries into either open or closed arm, and an increase in the proportion of time spent in the open arms relative to total spending time in either open or closed arm. Total arm entries were also used as indicators of changes in locomotor activities of the rats.
Hole-board test
The hole-board test was conducted according to the method of Takeda et al. [3]. The hole-board apparatus, raised to a height of 7 cm above floor level, was made of iron plate covered with dark Formica (62×62×36 cm) with 16 holes, each 4 cm in diameter, equally spaced in the floor. For the hole-board experiments, each rats was placed in the center of the hole-board and allowed to freely explore the apparatus for 5 min. The number of head dips and the time spent head dipping were recorded by a video camera for later analysis. A head dip was scored when the head was introduced into the holes at least to the level of the eyes.
Open field test
The rats were individually housed in a rectangular container that was made of dark polyethylene (60×60×30 cm) to provide best contrast to the white rats in a dimly lit room equipped with a video camera above the center of the room, and their locomotor activities (animal's movements) were then measured. The locomotor activity indicated by the speed and the distance of movements was monitored by a computerized video-tracking system using S-MART program (PanLab Co., Barcelona, Spain). After 5 min adaptation, the distance they traveled in the container was recorded for another 5 min. The locomotor activity was measured in centimeters. The time and number of line crossing (with all four paws) between the squares area was recorded for 5 min.
CORT analysis
Animals were killed by decapitation one day after behavioral measurement. For this, the unanesthetized rats were rapidly decapitated, and blood was quickly collected via the abdominal aorta. The blood samples were centrifuged at 4000 g for 10 min, and serum was collected and stored at -20℃ until use. The CORT concentration was measured by a competitive enzyme-linked immunoassay (ELISA) using a rabbit polyclonal CORT antibody (Novus Biologicals Corticosterone kit; Novus Biologicals, LLC., Littleton, CO, USA) according to the manufacturer's protocol. Samples (or standard) and conjugate were added to each well, and the plate was incubated for 1 h at room temperature without blocking. After wells were washed several times with buffers and proper color developed, the optical density was measured at 450 nm using an ELISA reader (MutiRead 400; Authos Co., Vienna, Austria).
Immunohistochemistry
For immunohistochemical studies, the primary antibodies against the following specific antigen were used: CRF (goat polyclonal CRF, 1:500 dilution, Santa Cruz Biotechnology Inc., California, CA, USA) and TH (sheep polyclonal TH, 1:2000 dilution, Chemicon International Inc., Temecular, CA, USA). Briefly, the sections were incubated with primary antiserum in PBST (PBS plus 0.3% Triton X-100) for 72 h at 4℃. The sections were incubated for 120 min at room temperature with secondary antibody. The secondary antibodies were obtained from Vector Laboratories Co. (Burlingame, CA, USA) and diluted 1:200 in PBST containing 2% normal serum. To visualize immunoreactivity, the sections were incubated for 90 min in avidin-biotin--peroxidase complex (ABC) reagent (Vectastain Elite ABC kit; Vector Labs. Co., Burlingame, CA, USA), and incubated in a solution containing 3,3'-diaminobenzidine (DAB; Sigma-Aldrich Chemical Co., St. Louis, MO, USA) and 0.01% H2O2 for 1 min. Finally, the tissues were washed in PBS, followed by a brief rinse in distilled water, and mounted individually onto slides. Images were captured using the DP2-BSW imaging system (Olympus, CA, USA) and processed using Adobe Photoshop (Adobe Systems, Inc., San Jose, CA, USA). The sections were viewed at 100× magnification, and the numbers of CRF and TH labeled cells was quantified in the hypothalamus and locus coeruleus (LC). CRF-, and TH-labeled cells were counted by an observer blinded to the experimental groups. Counting the immunopositive cells were performed within the square (200×200 µm2), anatomically localized in at least three different sections per rat brain according to the stereotactic rat brain atlas of Paxinos and Watson [22]. The stained cells for which intensities to a defined value above the background were only considered as immunopositive cells. The differences of brightness and contrast among raw images were not adjusted, in order to exclude any possibility of subjective selection of the immunoreactive cells.
Total RNA preparation and RT-PCR analysis
The expression levels of BDNF and TrkB mRNAs were determined by the reverse transcription-polymerase chain reaction (RT-PCR). The brain hippocampus was isolated from three rats per group. After decapitation, the brain was quickly removed and stored at -80℃ until use. The total RNA was prepared from the brain tissue using a TRIzol® reagent (Invitrogen Co., Carlsbad, CA, USA) according to the supplier's instruction. Complementary DNA was first synthesized from total RNA using reverse transcriptase (Takara Co., Shiga, Japan). PCR was performed using a PTC-100 programmable thermal controller (MJ Research, Inc., Watertown, MA, USA). The operating conditions were as follows: for glyceraldehydes-3-phosphate dehydrogenase (GAPDH), 30 cycles of denaturation at 95℃ for 30 sec, annealing at 58℃ for 30 sec, and extension at 72℃ for 30 sec; for BDNF, 27 cycles of denaturation at 95℃ for 30 sec, annealing at 57℃ for 30 sec, and extension at 72℃ for 30 sec; for TrkB, 38 cycles of denaturation at 95℃ for 30 sec, annealing at 59℃ for 30 sec, and extension at 72℃ for 30 sec. All primers were designed using published mRNA sequences of those cytokines and a primer designing software, Primer 3, offered by the Whitehead Institute for Biomedical Research (Cambridge, MA, USA; www.genome.wi.mit.edu) on the website. The following sequences were used: for GAPDH (409 bp), (forward) 5'-ATC CCA TCA CCA TCT TCC AG-3' and (reverse) 5'-CCT GCT TCA CCA CCT TCT TG-3'; for BDNF (153 bp), (forward) 5'-CAG GGG CAT AGA CAA AAG-3' and (reverse) 5'-CTT CCC CTT TTA ATG GTC-3'; for TrkB (347 bp), (forward) 5'-TGG GAC GTT GGG AAT TTG GTT-3' and (reverse) 5'-CAG CCG TGG TAC TCC GTG TG-3'. The PCR products were separated on 1.2% agarose gels and stained with ethidium bromide. The density of each band was quantified using an image-analyzing system (i-Max™, CoreBio System Co., Seoul, Korea). The expression levels were compared each other by calculating the relative density of target band, such as BDNF and TrkB, to that of GAPDH.
Statistical analysis
All measurements were performed by an independent investigator blinded to the experimental conditions. Results in figures are expressed as mean±standard error of means (SE). Differences within or between normally distributed data were analyzed by analysis of variance (ANOVA) using SPSS (Version 13.0; SPSS, Inc., Chicago, IL, USA) followed by Tukey's post-hoc test. Statistical significance was set at p<0.05.
RESULTS
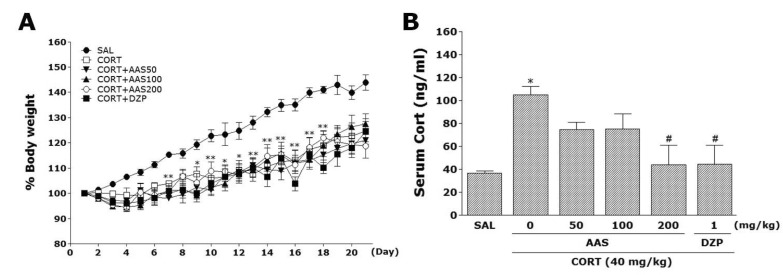
Effects of AAS on CORT-induced body weight loss and increase of serum CORT levels
Rats exposed to the repeated administration of exogenous CORT begin to lose body weight on the first day of CORT injections and this body weight loss is sustained for a prolonged period of time without restoration and is even exacerbated in some cases [23]. We measured the body weight of each rat in each group for 21 days. In the present study, body weight was evaluated daily for 21 days to identify whether the repeated administration of CORT (CORT group) would result in body weight loss (difference between daily weights and starting weight; Fig. 1A). Analysis of the body weight values revealed a significant gradual reduction of body weight gain over 21 days in the CORT group relative to control rats (SAL group). Interestingly, the body weight of rats treated with AAS showed no significant differences compared to that of rats in the CORT group during 21 days.
Additionally, the serum CORT levels were measured in each group following the repeated administration of CORT for 21 days. ELISA analysis revealed that CORT administration over 21 days significantly increased serum CORT concentrations by 284.81 % compared to saline-treated rats (Fig. 1B; p<0.05). This indicated that repeated CORT injections were sufficiently stressful despite the evoked CORT response (physiological response) to repeated CORT injections being significantly greater than the response to a single CORT injection (data not shown). In these results, the exogenous CORT-induced anxiety-like symptoms were exploited to develop a chronic stress model in rats. Daily administration of AAS significantly inhibited the exogenous CORT-induced increase of serum CORT levels compared to the CORT group (p<0.05).
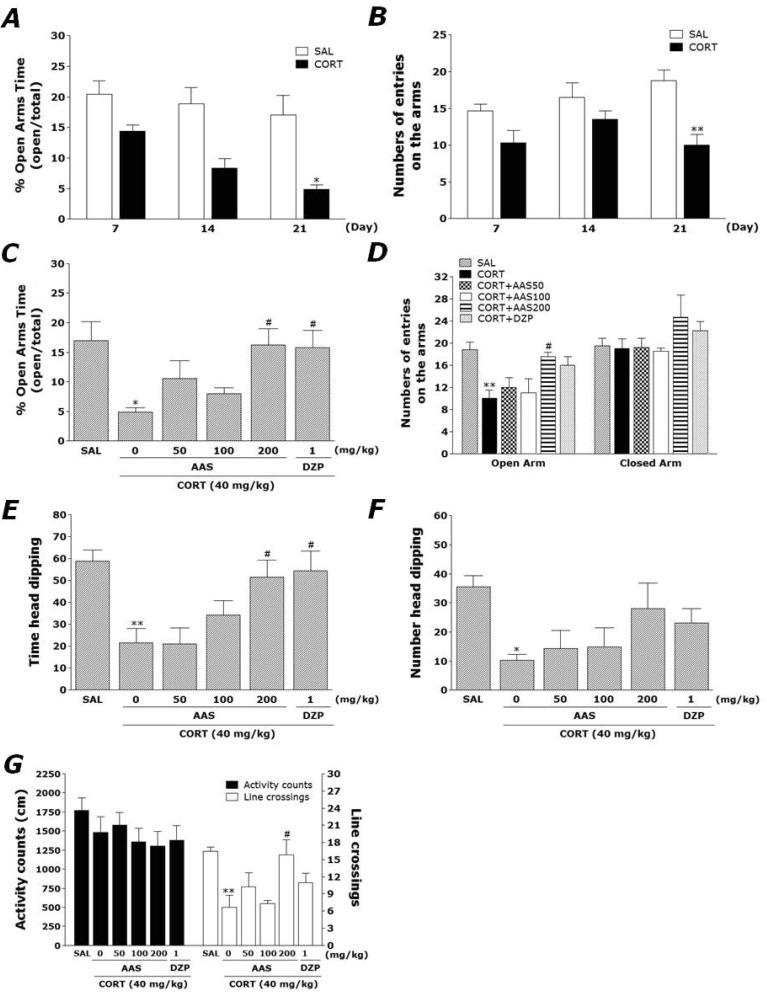
Effects of AAS on CORT-induced anxiety-like behaviors
Rats exhibited a marked anxiety phenotype characterized by decreased open-arm exploration during the EPM test. Rats were subjected to the EPM test 21 days after repeated CORT or saline injection (Fig. 2A and B). Immediately after repeated CORT administration (days 7 and 14), an decrease in both the percentage of time spent and the number of entries into the open arms of the maze were observed in the experimental group as compared with the SAL group. Furthermore, on day 21 from repeated CORT administration, rats displayed showed a significant decrease in both the percentage of time spent (p<0.05) and the number of entries (p<0.01) into the open arms of the maze compared with the SAL group. Conversely, statistical analysis revealed that the number of entries into the closed arms was not effected by 21 days from repeated CORT exposure, as there were no significant differences among the groups (data not shown).
The effects of AAS administration on anxiety-like behaviors, characterized by decreases in open-arm exploration in the EPM test, were also investigated. Post hoc comparisons revealed a significant decrease in the percentage of time spent by rats in the open arms of the maze following the repeated administration of exogenous CORT for 21 days compared to the saline-treated rats (p<0.05; Fig. 3A). However, the rats in the CORT+AAS200 group showed a significant restoration of the percentage of time spent, formerly decreased by repeated CORT injection, in open-arm of the maze, as compared to that in the CORT group (p<0.05). Similarly, post hoc comparisons revealed a significant decrease in the number of entries into the open arms of the maze after the repeated administration of exogenous CORT for 21 days compared to the SAL group (p<0.01; Fig. 2C). Rats in the CORT+AAS200 group also exhibited a significant restoration in the number of entries into the open arms of the maze compared to the CORT group (p<0.05). The lowest dose (50 mg/kg) decreased the percentage of entries in the open arms but did not change the percentage of entries in the closed arms. The highest dose (200 mg/kg) increased total arm entries, suggesting increased locomotor activity in the EPM test. Because no significant differences appeared in the number of closed-arm entries between groups in the EPM test, the observed anxiety-like behaviors of the rats receiving repeated CORT injections are likely not attributable to differences in their locomotor activities (Fig. 2D). AAS administration prior repeated administration of exogenous CORT did elicit anxiolytic or anxiogenic behavior in this study. These results reveal that the increase in the percentage of time spent into the open arms of the maze by the CORT+AAS200 group was almost comparable to those of the CORT+DZP group.
Effects of AAS on CORT-induced exploratory behaviors
The hole-board test been used widely to assess anxiety, emotionality or responses to stress in the animals [13]. An increase in head-dipping behaviors in the hold-board test reflects an anxiolytic-like state [1]. In the present study, 200 mg/kg AAS significantly increased time spent of head dips (p<0.05). Diazepam (1 mg/kg) also produced a significant increase in head dipping, in agreement with a previous study [13] (Fig. 2E and F). However, AAS-treated rats (200 mg/kg) displayed a significant increase in the total spent of head dips compared to the CORT group, indicating that anxiety-like behaviors in the CORT+AAS200 group was almost comparable to those of the CORT+DZP group. The data from the present study indicated that 200 mg/kg AAS has a significant anxiolytic-like effect in this paradigm.
Effects of AAS on CORT-induced motor functions or exploratory behaviors
Open-field activity was used to evaluate locomotor activity and exploratory behavior among the rats receiving CORT injections for 21 days (Fig. 2G). No significant differences appeared in locomotor activity (motor function) in the OFT among groups. However, rats receiving CORT injections displayed a significant decrease in the total number of line crossings compared to the SAL group (p<0.01). This finding suggested that CORT-treated rats subsequently produce exploration activities that are closely associated with anxiety-like behaviors in the OFT. However, AAS-treated rats (200 mg/kg) displayed a significant increase in the total number of line crossings compared to the CORT group (p<0.05).
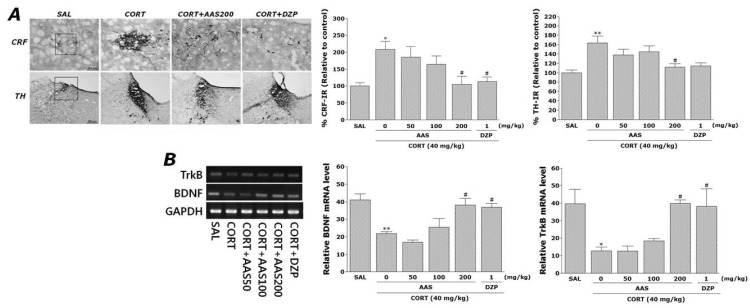
Effects of AAS on CORT-induced CRF- and TH-like immunoreactivities
Following the behavioral tasks, CRF-like immunoreactivity was analyzed in the cell bodies of various hypothalamic regions including the paraventricular nucleus (PVN; Fig. 3A). The numbers of CRF-immunoreactive fibers in the PVN of the CORT group were increased by 208.18%. Analysis of the numbers of CRF-immunoreactive neurons values revealed that rats receiving repeated administration of exogenous CORT exhibited a significant increase of CRF expression compared to the SAL group (p<0.05). The number of CRF-immunoreactive neurons was significantly decreased in the PVN region of the CORT+AAS200 group compared to the CORT group (p<0.05). This finding suggested that the increased CRF-immunoreactivity induced by the repeated administration of exogenous CORT was significantly restored by AAS administration and that the number of CRF-immunopositive neurons in the CORT+AAS200 group was closely associated with that in the CORT+DZP group (p<0.05). TH-like immunoreactivity was also analyzed in the locus coeruleus (LC) (Fig. 3A). The numbers of TH-immunoreactive fibers in the LC of the CORT group increased to 163.20%. Analysis of the numbers of TH-immunoreactive neurons values revealed that rats repeatedly exposed to exogenous CORT exhibit a significant increase of TH expression compared to the SAL group (p<0.01). The number of TH-immunoreactive neurons significantly decreased in LC of the CORT+AAS200 group relative to the CORT group (p<0.05). This finding indicated that the increase in the number of TH-immunoreactive neurons in rats repeatedly treated with exogenous CORT was significantly restored by AAS administration.
Effect of AAS on CORT-induced expression of BDNF and TrkB mRNAs in the hippocampus
The effect of AAS administration on the expression level of BDNF and TrkB mRNAs in rats with repeated CORT-induced hippocampus lesions were investigated using RT-PCR analysis (Fig. 3B). The BDNF mRNA expression levels were normalized against glyceraldehydes-3-phophate dehydrogenase (GAPDH) mRNA, an internal control. BDNF mRNA expression in the hippocampus in the CORT group was significantly decreased compared with that in the SAL group (p<0.01). The decreased expression of BDNF mRNA in the CORT group was significantly restored in the CORT+AAS200 group (p<0.05), and the restored level was similar to that of normal rats in the SAL group. Hippocampal expression of TrkB mRNA in the CORT group was also significantly decreased compared with that in the SAL group (p<0.05). The decreased expression of TrkB mRNA in the CORT group was significantly increased in the CORT+AAS200 group (p<0.05). This also indicated that the expression of BDNF and TrkB mRNAs in the hippocampus in rats receiving 200 mg/kg AAS administration was similar to that in rats receiving 1 mg/kg DZP administration.
DISCUSSION
AAS is a major compound isolated from the Korean medicinal herb Acorus gramineus, and it has multiple pharmacological activities, including antioxidant, anti-inflammatory, antitumor, antiviral, and antiapoptotic activities [14,16]. Recently, several studies have shown that AAS inhibits the activity of hippocampal neurons and produces an antiepileptic effect in the CNS by enhancing tonic GABAergic inhibition [24]. Other studies have also reported that β-asarone can attenuate neuronal apoptosis in rat hippocampus and cognitive impairment associated with conditions, such Alzheimer's disease [25,26], and can reduce the toxicity of excitatory amino acids and increase the expression of c-fos in the epileptic rat brain [27]. AAS has been shown to cross the blood-brain barrier and may act directly in brain nuclei to produce pharmacological effects in vitro and in vivo [28]. Thus, a new medicinal role is possible for AAS due to its anxiolytic action or anti-stress capabilities in the CNS, which may lead to the development of novel therapeutics for treating neurodegenerative or psychosomatic diseases in humans. Nevertheless, the reported pharmacological effects of AAS with respect to the anxiety-related symptoms seen in rats repeatedly exposed to exogenous CORT, as well as the mechanisms underlying these effects, remain unclear. Therefore, the purpose of the current experiment was to evaluate the ability of AAS to heal chronic stress-induced anxiety-like behavior in a rat model.
Our results demonstrated that repeated administration of AAS prior to CORT injection significantly increased open-arm exploration in the EPM and increased head dipping activity duration in the HBT. It seems likely that these behavioral effects are based on modulation of hypothalamic CRF activity, modification of the central noradrenergic system, and prevention of reductions in BDNF and TrkB expression in the hippocampus that underlie the development of anxiety. Thus, the current results support the possibility that AAS has anxiolytic effects.
Additionally, after 21 days of CORT treatment, a significant decrease in open-arm exploration time in the EPM test was observed compared with saline-treated controls. However, only small differences in the EPM behavioral test were observed on days 7 and 14 after terminating repeated CORT administration. Thus, 21 days of repeated CORT administration was found to be the optimal time to observe changes in anxiety-like behavior. Based on these results, we conclude that AAS pretreatment prior to each CORT treatment had anxiolytic effects, and that AAS may be a candidate therapeutic agent against psychosomatic diseases. Experimental animal models have become an invaluable and rapid tool in the analysis of a multitude of diseases: environmental or pharmacological agents can bring about symptoms, homologous to those of human anxiety disorders [8,23].
Furthermore, when measured immediately after behavioral testing, a gradual decrease in body weight gain and an increase in serum CORT levels were observed, indicating that the chronic CORT injections were sufficiently stressful [21,29]. Many studies have shown that chronic administration of high-dose CORT increased serum CORT concentrations in rats, in line with chronic stress models [30]. Accordingly, in animal models, forced sustainment of high CORT levels can affect anxiety-like symptoms under experimental conditions, and this might be related to the progression or exacerbation of chronically stressful conditions in humans [30]. The administration of AAS produced anxiolytic activity during the late period of AAS administration, suggesting that this therapy inhibited HPA axis-associated psychological dysfunction induced by decreased serum CORT levels. Thus, administration of AAS may modulate HPA axis dysregulation, indicating that it can influence endogenous CORT levels in the CNS, thereby normalizing behavioral and neurochemical responses. Similarly, serum CORT levels in rats treated with diazepam prior to performing the EPM test and HBT were lower than those of the controls, consistent with previous studies [3].
The results of these behavioral investigations revealed anxiolytic-like effects of AAS in animal models of anxiety. Many studies have suggested that stressed rats show decreases in the proportion of time spent and the number of entries made into the open arms in the EPM [31,32]. Although the EPM test is based on aversive contexts, conflicts, and the subsequent movement of the animal between an open and illuminated environment, the test includes two additional anxiety-provoking environmental parameters; height and open area [33]. In this test, it is assumed that animals feel safe in the closed arms but feel fear and anxiety during exploration of the open arms. In the present study, the administration of higher doses of AAS (200 mg/kg) prior to chronic CORT injections reduced anxiety-like behaviors significantly in the EPM test, as indicated by an increase in the number of entries into the open arms. Additionally, diazepam showed potential anxiolytic effects. However, in this study AAS at lower doses, 50 and 100 mg/kg showed no anxiolytic effect. Thus, these results suggest that the decreased anxiety-related behaviors following administration of AAS may be related to attenuated activity of the HPA axis.
We also investigated the effect of AAS in HBT, another murine test widely used to assess anxiety responses to an unfamiliar environment [34]. In the HBT, a pronounced inhibition of head-dipping behavior was observed in rats that had been exposed to stress stimuli [1]. Some studies previously reported that treatment with benzodiazepine anxiogenics or exposure to acute restraint stress produced a decrease in head-dipping behavior in mice [35]. These findings indicate that head-dipping behavior in the HBT is a good index for evaluating the emotionality of rodents. Our results indicated that higher doses of AAS (200 mg/kg) increased the time of head dipping behavior significantly, consistent with previous studies [3]. The dose-dependent activity of AAS (50, 100, or 200 mg/kg) was examined, and a dose of 200 mg/kg was the most effective in inhibiting chronic CORT-induced harmful effects in EPM test and HBT, which included anxiety-like behavior. The optimum dose determined in this study has been shown in a previous study [13].
An OFT was also performed to rule out any confounding motor impairments that can influence outcomes in behavioral test of anxiety [36]. No significant individual differences in locomotor activity were observed between groups, suggesting that the administration of AAS had no effect on sensorimotor performance. However, the administration of AAS prior to chronic CORT injections significantly reduces anxiety-like behavior, as indicated by an increase in total number of line crossing in the OFT. Accordingly, these results suggest that the observed increase of total number of line crossing in OFT is similar to observed changes in behavioral performances in the EPM test and HBT, which these results were likely due to anxiolytic activity.
The neuropeptide CRF plays an important role in the central regulation of stress and anxiety. In addition to its well-characterized ability to activate the HPA axis, CRF functions as a neurotransmitter at extrahypothalamic brain sites to mediate physiological and behavioral responses to stress [37]. Previous studies have shown that i.c.v. injections of CRF induced anxiety-like behavior, and increased plasma CORT via regulation of HPA axis hyperactivity [38]. The current data suggest that the CRF circuits in the PVN of the hypothalamus are activated by chronic CORT injections, leading to the observed anxiety-like activity in the behavioral tests [7]. These results show that the administration of AAS significantly blocked the increase in CRF immunoreactivity in the PVN. This suggests that anxiolytic effects following the administration of AAS are closely associated with CRF modulation in the PVN in the hypothalamus and activation of the HPA axis.
In addition, the enzyme TH is involved in stress-induced activation of the CNS and in stress-related psychopathological conditions such as anxiety [39,40]. These results are consistent with previous studies indicating that anxiety-like behavior induced by chronic stress is the result of changes in the CNS [41]. Moreover, we demonstrated that administration of AAS decreased TH-like immunoreactivity significantly in the LC of rats subjected to repeated CORT injection. Together, these findings suggest that AAS attenuated the behaviors and neurochemical responses associated with anxiety by modulating the HPA axis and the CNS, suggesting that administration of AAS, like diazepam, may indirectly alter catecholamine synthesis in the brain to produce physiological effects [13]. Thus, our results indicate that AAS acts by stimulating the CNS, suggesting that a dysregulated HPA axis may contribute to anxiety symptomology, and that the therapeutic action of the anxiolytic reverses this activity by increasing TH expression in the LC [42].
BDNF, an important neurotrophic factor, has also been implicated in the etiology of anxiety and the mechanism of anxiolytic treatment [43]. A clinical study demonstrated that stress-related psychiatric disorders including anxiety, are associated with reduced brain BDNF and its receptor TrkB levels [20,44]. In a recent experiment, chronic social defeat stress or treatment with exogenous CORT caused a significant decrease in BDNF expression in the hippocampus, a brain region that may be related to the pathogenesis of anxiety-like symptoms [20,45]. Consistent with these findings, we found the 3-week CORT injection decreased expression of BDNF and TrkB mRNAs in the rat hippocampus and anxiety-like behavior [20]. Decreased BDNF availability or reduced levels of the TrkB neurotrophin receptor could also reduce BDNF signaling [46]. However, administration of AAS restored the level of BDNF and TrkB mRNAs in the hippocampus of rats subjected to repeated CORT injection, suggesting that modulation of the BDNF-TrkB neurotrophic signaling pathway may play a role in mediating anxiolytic actions of AAS.
In summary, we demonstrated that the administration of AAS exhibited anxiolytic-like profiles in the EPM test and in the HBT, possibly via modifications of the central noradrenergic system and BDNF function. These findings indicated that AAS is capable of ameliorating the complex behaviors and neurochemical responses involved in anxiety by modulating HPA activity. Future studies need to focus on the mechanism of action of AAS involved in these anxiolytic properties.
 XML Download
XML Download