

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Asthma is a chronic inflammation characterized by airway hyperresponsiveness, airway narrowing, mucus overproduction, and airway remodeling. Airway inflammation via secretion of pro-inflammatory cytokines is essential for asthma development. These cytokines orchestrate allergic immune responses including T cell differentiation, IgE production, and leukocyte activation.12 Traditionally, allergic inflammation has been considered the result of environmental allergens such as dust mites, pollens, and animal dander. However, recent studies propose that bacteria-derived toxins or components can also be involved in allergic responses. Little is known about the mechanism by which environmental microbiota might contribute to allergic reactions.
A recent metagenomic data analysis revealed a microbiome in the respiratory tract, which was once believed to be sterile, mainly because of difficulties in culturing of the bacteria.3 Yet the airway is external to the body, and this space is thus a primary site for exposure to environmental factors such as bacteria and viruses. Differing compositions of lung microbiota between asthmatic and healthy people imply that bacteria might contribute to the development of disease.45 It is suggested that the local microbiota can directly affect airway-resident cells. However, the microbiota residing at other sites such as the gut might also affect lung immune responses. Further studies might elucidate interactions between the microbiota and the immune system that contribute to asthma.
An effective immune response requires appropriate cytokines and other inflammatory mediators produced by infected or resident cells. Microbiota- and host-derived molecules such as extracellular vesicles (EVs) might also regulate inflammation. EVs are lipid bilayers containing transmembrane proteins, cytosolic proteins, lipids, and nucleic acids with a diverse range of sizes (100–1,000 nm in diameter). As these molecules have surface ligands that interact with receptors on target cells, they can attach and modify the physiological state of recipient cells.67 Recent studies have demonstrated that EVs are involved in development of cancer, atherosclerosis, diabetes, meningitis, and salpingitis.
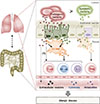
This review focuses on the potential relationship between environmental microbiota and asthma. We are of the view that asthma is not simply induced by allergens. We also propose that EVs are key molecules that link the microbiota to asthma by regulating immune responses.
Microbiota and immune responses of asthma
Asthma is a worldwide disease commonly characterized by eosinophilic inflammation. Given the large number of eosinophils in the airways, asthma is considered a hallmark of T helper type 2 (Th2) disorders of the lungs. The Th2 response is generally driven by the cytokines IL-4, IL-5, IL-9, and IL-13, which promote increased numbers of eosinophils in the airway and lead to high IgE levels in the blood. In mice, depletion of Th2-associated cytokines or induction of the Th1 response has reduced asthma features.89 However, there is an increasing interest in understanding the non-Th2 response, which likely represents a large proportion of asthma cases.
The non-Th2 response involves adult-onset and severe asthma, which have a mixed Th1 and Th17 response with neutrophilic inflammation.1011 In mice models of asthma, IL-17 has been strongly linked to neutrophilic inflammation, and has led to corticosteroid resistance.1213 Severe asthma driven by IL-17 pathologically correlated with steroid-resistant asthma. Neutrophilic asthma has been linked to upregulation in TNF-α pathways, but it remains uncertain whether TNF blockade could improve steroid responsiveness.1415 Although 2 forms of asthma have been defined, Th2 and non-Th2 responses co-occur in many cases, rather than being mutually exclusive.
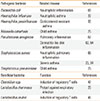
Similar to many chronic inflammatory diseases, the pathogenesis of asthma mainly depends on genetic susceptibility and environmental factors.161718 Environmental changes are a major factor in the development of allergic diseases. The recent microbiota hypothesis has suggested that perturbations in the gut microbiota disrupt the mucosal tolerance.1920 A balance between the microbiota and host that maintains functional homeostasis is accurately regulated; however, it is constantly challenged by several factors. Once pathogenic bacteria colonize a niche, they cause a state of dysbiosis in the microbial community, which is now recognized to induce allergic diseases such as asthma (Table).
Airway microbiota and asthma
The presence of certain bacteria in the airway can regulate allergic inflammation.21 Haemophilus influenzae might be related to neutrophilic asthma, in which inflammation is mainly induced by neutrophils.22 Another well-known pathogenic bacterium, Staphylococcus aureus, is thought to be involved in severe asthma, since asthmatic patients have presented high levels of specific antibodies against S. aureus enterotoxins.2324 Other bacteria such as Moraxella catarrhalis and Streptococcus pneumoniae have also been associated with childhood asthma.25 However, not all bacteria in the airway aggravate the severity of asthma; for example, Lactobacillus rhamnosus has protective effects against respiratory infection.26 Specific bacteria seem to modulate allergic inflammation; however, whether a single bacterial species can induce allergic disease is still unclear.
Recent studies have revealed that the composition of the microbiota residing in the lung can be more important in allergic diseases than the simple presence of individual species. Analyses of 16S rRNA have found that the airway hosts a complex community of microbes; moreover, bacterial populations of asthmatic patients are unlike those of healthy control subjects.2728 The bacterial compositions of bronchial samples have also exhibited differences between 2 groups.2930 These reports indicate that the phylum Proteobacteria was relatively prevalent in asthmatics, while phylum Bacteroidetes predominated in healthy controls. It has become clear that microbial community dysbiosis correlates with asthma.3132 Diverse bacterial communities reside in the airway, and altered composition of these microbiota might contribute to allergic inflammation in asthma.
Clinical studies of corticosteroid inhalation suggest that the airway microbiota affects corticosteroid responsiveness among asthmatic patients.33 Bacterial community profiles of corticosteroid-sensitive or -resistant asthmatics were not well discerned, but Haemophilus parainfluenzae was observed in some of the corticosteroid-resistant asthmatics. Another study of antibiotic treatment observed azithromycin-induced modification of the airway microbiota of adult asthmatic patients.34 The abundance of members of the Haemophilus and Pseudomonas genera declined within the community, but the abundance of Anaerococcus species increased. These findings suggest that the airway microbiota drives asthma development, and explain corticosteroid responsiveness. We expect that current efforts to understand the airway microbiome membership in detail will help reveal a mechanistic link between the microbiota and asthma.
Gut microbiota and asthma
The airway microbiota can shape lung-specific immune responses resulting in either homeostatic or detrimental inflammation. Recently, the gastrointestinal (GI) tract microbiome has also become a plausible influencer of immune development against allergy or asthma. This interest in GI microbes stemmed from an observed inverse relationship between rate of childhood asthma and exposure to bacteria in infancy.35363738 Moreover, infants who develop asthma have presented relatively few lactobacillus, and bifidobacteria, and relatively many enterococcus.394041 Further studies indicate that the gut microbiota influences the maturation of immune function in early life through the oral ingestion of environmental bacteria.424344
Studies in mice also provide strong evidence of the role of gut microbiota in the regulation of immune function. Germ-free (GF) mice presented strong allergic inflammation compared to specific pathogen-free (SPF) mice through OVA sensitization and challenge.45 Specific bacteria like Lactobacillus reuteri exhibited the protective effects of inducing regulatory T (Treg) cell expansion and reducing inflammation, in response to OVA challenge in sensitized BALB/c mice.46 Treatment with Clostridium strains produced similar effects on Treg cells in the colonic mucosa, and reduced IgE levels after OVA sensitization.47 Interestingly, segmented filamentous bacteria induced Th17 cell expansion, which mediates mucosal defense in the lamina propria.48 However, a recent study found that direct ingestion of certain bacteria was insufficient to induce significant changes in immune function.
Experimental feeding of different dietary components indicated that dietary changes could affect allergic inflammation by modifying the microbiota composition. High fiber diet altered the ratio of Firmicutes spp. to Bacteroidetes spp., and increased the levels of circulating short-chain fatty acids. These subsequent alterations protected against allergic lung inflammation via activation of Treg cells.4950 The mechanisms of Treg-mediated suppression are not fully understood, but microbes do promote Treg cell activation by maintaining resident dendritic cells (DCs) in an immature state, which is essential for mucosal tolerance.5152
The propensity of an innocuous antigen to induce local and systemic immune unresponsiveness is termed as oral tolerance.53 In the large intestine, commensal bacteria are also regulated by an analogous, but more locally processed, tolerance. It is assumed that oral tolerance and airway tolerance are tightly linked, and that the GI tract might act as a sensor for the development of tolerance to antigens. However, it remains to be determined how changes in the gut microbiota affect lung immunity.
EVs as mediators of allergic disease
Immune system recognition of pathogens is essential to activating immune cells. Bacterial components that trigger immune response are often referred to as pathogen-associated molecular patterns (PAMPs); these components include carbohydrates, lipids, proteins, and genetic material. PAMPs can bind to pattern recognition receptors (PRRs) on immune and non-immune cells, thereby initiating signaling cascades. Given their role in the immune system, pathogens have evolved to increase their virulence by modulating their signals to PRRs.54 Recent studies report that both PAMPs and EVs released from infected cells and pathogens are likely to be involved in immune responses.55
EVs are released by diverse cell types that have been implicated in allergic responses such as bronchial epithelial cells, mast cells, dendritic cells, and T cells in the lung.56 Bronchial epithelial cells are the primary producers of EVs in the lungs of patients with asthma.57 Interestingly, it was found that IL-13-stimulated bronchial epithelial cells released EVs that promoted proliferation of macrophages, but reduction in EV secretion seemed to ameliorate asthmatic features. During allergic inflammation, mast cell-derived EVs were highly activated, and contributed to induction of DC maturation.58 DCs also produce EVs that present allergens and activate allergen-specific Th2 cells.59 Many studies suggest that EVs can transfer MHC/antigen complexes, enabling the DCs to efficiently activate T cells.6061 These observations indicate the potential contributions of diversely sourced EVs to the pathogenesis of asthma. In contrast, some studies have reported that EVs can inhibit Th2 response-associated cytokine production, IgE response, and even prevent the development of asthma.62 While immune cells mediate immune responses, non-immune cells such as bacteria also have important roles in activating immune responses. EVs produced by pathogens such as Staphylococcus aureus have been implicated in atopic dermatitis-like skin inflammation;6364 these EVs might also induce neutrophilic pulmonary inflammation.65 EVs derived from E. coli induce several infectious diseases, and can aggravate emphysema through IL-17A-mediated neutrophilic inflammation.66 Pseudomonas aeruginosa often contributes to lung diseases such as cystic fibrosis, and its EVs increase pulmonary inflammation through Toll-like receptor (TLR)2 and TLR4 pathways.67 Moreover, indoor dust, which contains many components of bacteria, has been associated with both Th1 and Th17 responses, which induce neutrophilic pulmonary inflammation.6869 As bacteria-derived EVs can affect distal host cell sites,70 these vesicles might play significant roles throughout the body. Here, we propose a model that uses EVs to link the environmental microbiota with airway immunity (Figure).
CONCLUSIONS
The presence of lung and gut dysbiosis in asthma suggests that microbiota composition might shape the immune system and lead to the development of asthma. Microbial communities have immunomodulatory roles in both the disease progression and the clinical outcome of asthma, but our understanding of mechanisms linking gut- to lung-immunity is still unclear. EVs might provide new insights into immune responses, as they can move to distant sites and stimulate other cells such as immune and epithelial cells. EVs produced by commensal bacteria can benefit the host by promoting mucosal tolerance and protecting against the onset of diseases. However, questions remain as to which components of EVs activate recipient cells and how EVs regulate intracellular signaling pathways. Further studies will provide a mechanistic understanding of interactions between the microbiota and the host immune response through EVs, which in turn will aid in providing new insights into the management of asthma.
 XML Download
XML Download