

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Asthma is a chronic inflammatory disease that is characterized by infiltration of inflammatory cells such as eosinophils, airway hypersensitivity, and airflow obstruction.1,2 Th2 cells play a particularly crucial role in the development of asthma by secreting interleukin (IL)-4, IL-5, and IL-13: IL-4 causes naïve T cells to differentiate into Th2 cells while IL-5 induces eosinophil infiltration.3
The hygiene hypothesis proposes that westernized culture and its associated improvements in sanitation cause the microbial exposure of the population to be reduced. This decreases Th1 immune responses that are induced, the lack of which causes the immune system to become dominated by Th2 immune responses.4,5,6 It has been suggested that this skewing of the immune system has caused the incidence of allergic diseases such as asthma to rise rapidly in the last few decades.7,8,9 However, recent studies suggest that the incidence of Th1-related diseases such as type 1 diabetes mellitus and multiple sclerosis is also increasing along with Th2-related diseases such as asthma.10 This indicates that other mechanisms apart from Th1/Th2 immune responses participate in the modulation of allergic inflammatory responses. One of these mechanisms may involve regulatory T cells (Tregs): several studies report that Tregs regulate immune system by suppression of the Th1/Th2 responses.11,12,13 Tregs, which are widely considered to be crucial suppressor T cells, act by releasing the immunosuppressive cytokines IL-10 and transforming growth factor (TGF)-β14 and by cell-cell contact mechanism that is mediated cytotoxic lymphocyte associated antigen-4 (CTLA-4) expressed by CD4+CD25+ Tregs, which interacts with CD80 and/or CD86 on the surface of antigen-presenting cells (APCs).15
Bacillus Calmette-Guérin (BCG) is a mycobacterial vaccine organism that is used to prevent tuberculosis. It is known to modulate allergic inflammatory responses.16,17 For example, infants with a BCG scar have a higher anti-inflammatory response, namely their ratio of IL-10-producing T cells to IL-4-producing T cells is higher than the ratio of controls who lack a BCG scar.18 Significantly, analyses of mice who were immunised with BCG in the neonatal period revealed that their splenocytes had a higher frequency of CD4+CD25+ Tregs, expressed more Foxp3 and CTLA-4, and produced more IL-10 and TGF-β.19 Thus, BCG may be an example of the microbial exposure that is proposed in the hygiene hypothesis to prevent susceptibility to allergic inflammation.
Dendritic cells (DCs) are APCs that play an important role in modulating immune responses.20,21,22,23 DCs can be divided into immature and mature DCs. Contact with antigens can cause immature DCs to differentiate into mature DCs that express on their cell surfaces CD80, CD86, CD40, and major histocompatibility (MHC) class II markers.
Many studies have reported that depending on the stimuli, mature DCs can prime naïve T cells toward Tregs. Mature bone marrow-derived DCs (BMDCs) were able to induce the proliferation of CD4+CD25+ T cells in the presence of a polyclonal stimulus and in the absence of exogenous IL-2.24 Thymic stromal lymphopoietin-treated thymic DCs express CD80/86 and MHC class II and promote the conversion of CD4+CD25- thymocytes into CD4+CD25+Foxp3+ Tregs.25 Mature human monocytes-derived DCs upregulate the immune-inhibitory enzyme, indoleamine 2, 3-dioxygenase and expand CD4+CD25bright Foxp3+CD127neg Tregs.26
We confirmed previously that vaccination with BCG can suppress asthma-related inflammatory responses in a mouse model of asthma.27 The aim of the present study was to investigate the mechanism by which BCG vaccination inhibits the inflammatory allergic responses of this mouse model; to do so, the roles of DCs and Tregs were analysed.
MATERIALS AND METHODS
Animals
Female BALB/c mice were purchased from Orient Bio (Seongnam, Korea) and cared for and used in accordance with the guidelines of the Institutional Animal Care and Use Committee (IACUC) at Asan Medical Center and Ulsan University College of Medicine. The mice were housed in specific pathogen-free facility.
Antigen sensitization and challenge
The mice (6 weeks of age, 5 mice/group) were sensitized by 2 intraperitoneal injections with 10 µg ovalbumin (OVA; chicken egg albumin grade V, Sigma Chemical Co., St. Louis, USA) dissolved in 100 µL of phosphate-buffered saline (PBS) adsorbed to an equal volume of alum solution (Alumimject, Pierce, IL, USA) on days 0 and 7. On days 14-16, all mice were challenged daily for 30 minutes via an ultrasonic sprayer (Nescosonic UN-511, Alfresa, Osaka, Japan) with aerosolized 1% OVA (50 µg in 50 µL saline). The mice were divided into 5 groups of 5 mice each.
BCG vaccination
Freeze-dried living BCG vaccine was obtained from Japan BCG laboratory (Tokyo, Japan). The mice were injected intraperitoneally with BCG Tokyo 172 strain 5 days before the first sensitization (day -5) at a dose of 1×106 CFU/200 µL.
DCs preparation, BCG treatment, and injection into mice
BMDCs were generated from BALB/c (6-8-weeks-old). Briefly, bone marrow cells were cultured in Iscove's Modified Dulbecco's Medium (IMDM) supplemented with 10% fetal bovine serum (FBS), 1% L-glutamine, antibiotic-antimycotic, and 10 ng/mL recombinant murine Granulocyte-macrophage colony-stimulating factor (GM-CSF) (R&D systems, Minneapolis, MN, USA). The culture media were replaced with fresh media containing GM-CSF on days 3 and 6. On day 6, BMDCs were collected and stained for CD11c. then, the CD11c+ cells were sorted by using autoMACS (Miltenyi Biotec) and prepared at 1×106 cells/10 mL, and stimulated with BCG (1×107 CFU) for 48 hours in vitro. The mice were injected intravenously with BCG-stimulated DCs or non-stimulated DCs (1×106/200 µL) 5 days before the first sensitization (day -5).
In vivo treatment with anti-CD25 monoclonal antibody
The administration of anti-CD25 Abs is a commonly used method to deplete Tregs in vivo.28 To investigate the mechanism by which Tregs participate in the allergy-suppressive effects of vaccination with BCG or BCG-stimulated DCs, the OVA-sensitized mice were injected intraperitoneally with 250 µg of anti-CD25 monoclonal antibody (mAb) (clone PC61, eBioscience, San Diego, USA) 1 day before the first challenge with 1% OVA.
Measurement of bronchial hyperresponsiveness (BHR)
Twenty-four hours after the final OVA challenge, BHR was measured in conscious unrestrained mice by using a barometric whole-body plethysmograph (one chamber plethysmography, All Medicus, Anyang, Korea), as described previously.29 Briefly, after inhalation of saline and each concentration of methacholine (MeCh), enhanced pause (Penh) was measured at 10-seconds intervals for 3 minutes, averaged, and expressed as absolute values.
Bronchoalveolar lavage (BAL) and determination of the cellular composition of the BAL fluid
After measurement of BHR, mice were anaesthetized. BAL was performed as described previously.29 Briefly, the trachea was immediately exposed after anesthesia. Then, BAL was performed through a catheter inserted into the exposed trachea following instillation of normal saline (2 mL) at 37℃. The BAL fluid was centrifuged at 2,000 rpm for 5min at 4℃. After discarding the supernatant, the pellet was resuspended in 100 µL PBS. Total BAL cell counts were performed using a haemacytometer. To count the different BAL fluid cells, cytospin slides were prepared, stained with Wright stain, and the different cell types were identified on the basis of their standard morphology under a light microscope. Five sections per slide were evaluated.
Quantitation of the serum levels of immunoglobulins
Serum was collected upon sacrifice 24 hours after last 1% OVA challenge and the serum immunoglobulins were quantified by sandwich ELISA as described previously.29 Briefly, serum was separated from the blood clot by centrifugation at 2,500 rpm for 10 minutes at 4℃. Absorbance of Total IgE, OVA-specific IgE, IgG1 and IgG2a was measured using ELISA reader. The optical density was measured at 450 nm.
Histopathological analysis of the lungs
To histologically evaluate the lung tissues, lung samples were prepared as described previously.30 Inflammation was scored by 2 independent blinded investigators. The degree of peribronchial and perivascular inflammation was evaluated on a subjective scale of 0-3 as described elsewhere.30 Briefly, a value of 0 meant that no inflammation was detectable, a value of 1, occasional cuffing with inflammatory cells, a value of 2, most bronchi or vessels were surrounded by thin layer (1 to 5 cells thick) of inflammatory cells, and a value of 3, most bronchi or vessels were surrounded by a thick layer (more than 5 cells thick) of inflammatory cells. The cellular infiltration in 5 randomly selected fields was assessed under a Zeiss Axiophot microscope (Carl Zeiss Inc., Thornwood, USA) at 100×.
RESULTS
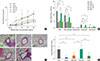
Transfer of BCG-stimulated DCs inhibits BHR and airway inflammatory cells
The MeCh-BHR in response to aerosolized MeCh was measured and the cells in the BAL fluid were identified and counted. Mean Penh value at maximum MeCh dose of the BCG-stimulated DCs mice was half those of mice, which had only been sensitized and challenged with OVA (OVA mice) (P<0.05, Fig. 1A). By contrast, the mean Penh values at maximum MeCh dose of the mice who had received non-stimulated DCs before OVA sensitization and challenge were equivalent to those of the OVA mice. In terms of the BAL fluid analyses, the mice receiving the BCG-stimulated DCs had significantly fewer infiltrating cells overall than the OVA mice (P<0.01, Fig. 1B), whereas the mice receiving non-stimulated DCs were similar to the OVA mice. With respect to specific cell types in the BAL fluid, the mice receiving the BCG-stimulated DCs had significantly fewer eosinophils than the OVA mice and the mice receiving non-stimulated DCs (P<0.01, Fig. 1B).
Transfer of BCG-stimulated DCs suppresses the inflammation of lung
Histological analysis of the lungs of the mice described above revealed that the recruitment to the lungs of peribronchial and perivascular inflammatory cells was low in mice who had received the BCG-stimulated DCs when compared to the OVA mice (Fig. 1C). In contrast, the recruitment of the mice that received the non-stimulated DCs was equivalent to that of the OVA mice (Fig. 1C).
To quantify the infiltration of inflammatory cells, peribronchial and perivascular histopathological inflammation scores were determined. Compared to the OVA mice, the inflammation score was low in the mice that had received BCG-stimulated DCs (P<0.01, Fig. 1D). By contrast, the scores of the mice that had received the non-stimulated DCs were similar to those of the OVA mice (Fig. 1D).
Transfer of BCG-stimulated DCs reduces the systemic immune response to OVA
The levels of total IgE, OVA-specific IgE, OVA-specific IgG1, and OVA-specific IgG2a in the sera of each group were measured. The OVA mice had elevated levels of total IgE and OVA-specific IgE. Both measures were significantly lower in the mice that received the BCG-stimulated DCs but not in the mice that received the non-stimulated DCs (P<0.05, P<0.01, respectively, Fig. 2). Compared to the OVA mice, the mice that received the BCG-stimulated DCs also tended to have lower OVA-specific IgG2a levels and their OVA-specific IgG1 levels were significantly lower (P<0.05, Fig. 2). By contrast, the OVA-specific IgG1 and OVA-specific IgG2a levels of the mice that received the non-stimulated DCs were similar to those of the OVA mice (Fig. 2).
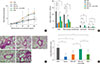
Anti-CD25 mAb treatment prior to OVA challenge can abrogate the effect of BCG-stimulated DCs transfer
BHR analysis revealed that the mean Penh values at maximum MeCh dose of the CD25-depleted mice were significantly higher than those of the BCG-DCs-injected mice (P<0.05, Fig 3A). BAL fluid analysis revealed that the anti-CD25 mAb treatment also significantly increase the total cell numbers and eosinophils relative to the mice that treated the BCG-stimulated DCs (P<0.01, respectively, Fig. 3B).
There was significantly more recruitment of inflammatory cells to the lungs in the anti-CD25 mAb treated mice than in the BCG-stimulated DCs-treated mice (P<0.01, Fig. 3C and D).
DISCUSSION
Many studies in human and experimental work in animals showed that BCG can modulate the allergic inflammatory responses.16,17,18 However, the mechanism(s) by which this occurs remained unclear. In the present study, we asked whether this mechanism is associated with Tregs that are stimulated by BCG-stimulated DCs.
The present study shows that BCG vaccination and the transfer of BCG-stimulated DCs both inhibited asthmatic responses, including BHR, serum IgE production, and the numbers of inflammatory cells in the BAL fluid and lungs. In addition, treatment with anti-CD25 mAb prior to challenge abrogated these effects. These findings suggest that BCG inhibits allergic inflammatory responses by inducing Tregs, which is mediated by DCs.
The first of these experiments involved injecting mice with BCG-stimulated DCs, as this would reveal whether the inhibitory effect of BCG is associated with DCs. Indeed, compared to the OVA mice and the mice that received non-stimulated DCs, the transfer of BCG-stimulated DCs significantly suppressed the asthmatic responses, including BHR, the numbers of inflammatory cells in the BAL fluid, and the pulmonary inflammation in the lung tissue. This effect of BCG has also been observed by other studies.31,32,33 These results suggest that the BCG-induced maturation of DCs plays an important role in regulating allergic immune responses.
Our second experiment involved the administration of anti-CD25 mAb to determine whether the inhibitory effect of the BCG-stimulated DCs is associated with Tregs. Cell-surface CD25 is a well-known marker for Tregs.34 Tregs depletion by CD25 mAb treatment indeed enhanced BHR and measures of airway inflammation, namely the total cell and eosinophil counts in the BAL fluid and the infiltration of inflammatory cells in the lung. Thus, it appears that the asthma-inhibiting effects of BCG are associated with Tregs. These observations are consistent with those of other studies that have used mouse models.35,36,37 For example, BCG-primed DCs were shown to enhance the expression of Foxp3+ Tregs, IL-10, and inducible costimulator.35 However, interestingly, BCG-primed DCs have also been shown to upregulate Th1-associated neutrophilic inflammation; this was more effective when the Pasteur strain was used instead of the Copenhagen strain.35 In the present study, the Tokyo strain of BCG was used and neutrophil increases were not observed. Thus, this difference may be related to the type of BCG strain that is used. Supporting this is a study that found considerable differences in the immunogenicity of various BCG strains, namely their ability to modulate T-cell proliferation and cytokine production.38
This study demonstrated that matured DCs by BCG in vitro could suppress allergic inflammation in mouse model of asthma with adoptive transfer. In light of these, our study provides a new perspective of the mechanisms by which the allergic inflammation in a mouse model of asthma can be modulated.
Further studies are needed to refine several aspects of this study. CD25 is not a specific marker of Tregs and it may be expressed by activated T and B cells. Therefore, the use of anti-CD25 may not be sufficient to explain the mechanism of Tregs.39 And we did not investigate the expression of Tregs in the airway or thymus or regional lymph node and the functional cytokine production. However, BCG treatment is likely to elevate Foxp3+ Tregs which is a master transcription factor of Tregs in vivo and in vitro.35,36 In addition, recent study has reported that DCs stimulated by BCG release IL-10 and induce differentiation of naïve T cells into Tregs.39
BCG is also recognized as a strong Th1 immune response inducer.41,42 A study has reported that the interaction between BCG and toll-like receptor (TLR) 2 on DCs may affect the differentiation of naïve T cell mainly to Th1-like cells with the help of IL-10, IL-12, and so forth.43 In addition, recently, some studies have reported that IFN-γ is essential to suppress allergic responses.44 It would be worth to perform the study of BCG-DCs on damping allergic inflammation in IFN-γ- or TLR2-deficient mice.
In summary, the present study showed that BCG treatment inhibited the allergic responses of a mouse model of asthma; the study with BCG-stimulated DCs was consistent with these observations. In addition, anti-CD25 mAb treatment reversed the effects of BCG/BCG-DCs. These results suggest that the inhibitory effect of BCG in our mouse model of asthma is mediated by DCs, which induce Tregs.

 XML Download
XML Download