

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Human enteroviruses (EVs) are positive-sense, single-stranded, RNA viruses and belong to Picornaviridae (1). Sequence analysis of human enteroviruses further grouped them into four species (EV-A to D). Among these, six serotypes of EV-A (CAV2, 4, 6, 10, 16 and EV71) and four serotypes of EV-B (E6, 9, 30 and CAV9) have been detected in young children with sepsis-like illness across Edinburgh and the US (234).
Echovirus 30 (E30) is a key pathogen related with aseptic meningitis in humans (56). It is highly prevalent on a global scale (789). The transmission routes of E30 may be direct such as fecal-oral and respiratory routes or indirect, such as via contaminated water (10).
Primary infection with E30 leads to replication of the virus in the tissue around the gastrointestinal tract, with transient viremia following (11). Infectious diseases caused by pathogenic organisms such as viruses are still responsible for the majority of hospitalization and death worldwide. Although considerable advancement has been made by using various antibiotic agents, there remain big challenges in the effective treatment of various infections (1213). In addition to numerous side effects in the use of antiviral agents due to their effect on the whole body, the rapid emergence of drug resistance adds a serious problem to human health (1415). Moreover, new viruses are constantly emerging through evolution or other biological events, leading to a continuing challenge in the control and prevention of infectious diseases. Consequently, in addition to discovering novel antiviral drugs for infection control, new strategies need to be expanded to maximize efficacy of the existing available drugs (16).
Gemcitabine is widely utilized in the treatment of pancreatic, ovarian, and non-small cell lung cancers (17). To date, however, no reports have described the antiviral activity of gemcitabine against E30. In this study, we assess the antiviral activity of gemcitabine against E30, and further elucidate its mechanism of action.
MATERIALS AND METHODS
Viruses and cell lines
E30 (ATCC, VR-692) was purchased from American Type Culture Collection (Manassas, VA, USA) and grown at 37℃ in RD cells (human rhabdomyosarcoma cell line). RD cells were cultured in minimal essential medium (MEM), supplemented with 10% fetal bovine serum (FBS) and 1% antibiotic-antimycotic solution. MEM, FBS, trypsin-EDTA, and antibiotic-antimycotic solution were purchased from Gibco BRL (Invitrogen Life Technologies, Karlsruhe, Germany) and tissue culture plates were purchased from Falcon (BD Biosciences, San Jose, CA, USA).
Antiviral activity assay
Antiviral activity was determined by sulforhodamine B (SRB) staining using the cytopathic effect (CPE) reduction assay method, following viral infection (18). One day before infection, RD cells (at 2×104 cells/well) were seeded onto a 96-well plate (Falcon, BD biosciences, San Jose, CA, USA). The next day, culture medium was altered to 1% FBS and the virus suspension was added at a 50% tissue culture infective dose (TCID50). An appropriate concentration of the gemcitabine was added to the plate which was then cultured at 37℃ in a CO2 incubator for 2 days until the appropriate CPE was generated. To fix cells, ice-cold 70% acetone was added to the plate. After 30 min, the fixed cells were stained using SRB solution (0.4% (w/v) in 1% acetic acid solution. To solubilize bound SRB, 10 mM unbuffered Tris base solution was added to the plate. The absorbance was measured at 562 nm by a SpectraMax i3 microplate reader (Molecular Devices, Palo Alto, CA, USA) with a reference absorbance at 650 nm. Cell viability (%) was calculated for comparison, based on the optical density measured. In addition, images of SRB-stained cells were recorded using ImmunoSpot (CTL, ShakerHeights, OH, USA).
To test the effect of gemcitabine on the infectivity of E30 particles, E30 was pre-incubated with gemcitabine or rupintrivir at 4℃. After 1 h, RD cells were treated with pretreated or untreated E30 at 37℃ and following a further 1h, unbound virus was eliminated. After washing using 2x PBS, RD cells were incubated in medium provided with or without gemcitabine or rupintrivir at 37℃. Antiviral activity was measured using the SRB assay 2 days following (19).
Time course experiment
RD cells infected with TCID50 E30 were harvested at various time points, including 4, 6, 8, 10, and 12 h post-infection, after which gemcitabine and rupintrivir were added. Total RNA was isolated at the indicated time points post-infection. The levels of E30 RNA were analyzed using real time-PCR.
Time-of-addition (TOA) assay
Gemcitabine and rupintrivir were then added onto the cells at 10 µM or 2 µM, respectively, either before (−1 h), during (0 h), or after (1, 2, 4, 6, 8, 10 and 12 h) E30 infection. After 14 h, real-time (RT−) PCR analysis was conducted using Thunderbird SYBR qPCR Mix (Toyobo, Osaka, Japan).
Real-time PCR
Total RNA was extracted using the QIAamp viral RNA Mini kit (Qiagen, Valencia, CA, USA). The reverse transcription reaction was generated using an RNA 20µl reaction volume for 60 min at 42℃, containing RNase inhibitor, murine Maloney leukemia virus reverse transcriptase with 5×buffer, oligo(dT) 15 primer and a dNTP mixture (all from Promega, Madison, WI, USA) according to the manufacturer's instructions. Quantitative RT-PCR analysis was conducted and cDNA was amplified using the CFX96 optical reaction real-time PCR system (Bio-Rad, Hercules, CA, USA) with Thunderbird SYBR qPCR Mix (Toyobo, Osaka, Japan). The following primers were used: E30 5′-NCR-sense, 5′-TCC TCC GGC CCC TGA ATG-3′ and 5′-NCR-antisense, 5′-GAA ACA CGG ACA CCC AAA G-3′ (20).
RESULTS
In vitro antiviral activity of gemcitabine against E30 infection
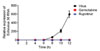
We recently reported that gemcitabine has antiviral activity against human rhinoviruses (21). Here we confirmed the antiviral effect of gemcitabine on E30 by assessing the virus-induced cytopathic effect in RD cells using an SRB assay. E30 induced approximately 90% cell death in RD cells, and treatment of cells with 2 µM of gemcitabine significantly increased cell viability up to 80%. Furthermore, 10 µM of gemcitabine fully preserved RD cells from cytotoxicity induced by E30 infection. Rupintrivir (positive control) exhibited strong antiviral activity against E30 up to a concentration of 0.016 µM (Fig. 1A). We also assessed the cytotoxicity of gemcitabine and rupintrivir against RD cells. Gemcitabine and rupintrivir did not show cytotoxicity at 50 µM and 2 µM respectively (Fig. 1B). Collectively, these results suggest that gemcitabine possesses antiviral activity against E30 in vitro.
Gemcitabine affects intracellular processes during early stages of E30 infection
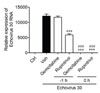
Based on the results produced from the E30 in vitro assay, we conducted time course experiments to analyze the mechanism of action of gemcitabine. Viral RNA from E30 was detected using RT-PCR at various times points (4, 6, 8, 10, and 12 h post-infection) and after gemcitabine (10 µM) and rupintrivir (2 µM) treatment of E30-infected cells. We could find E30 viral RNA as early as 8 h post-infection. Interestingly, gemcitabine was seen to block the proliferation of E30 in resemblance to rupintrivir (inhibitor of picornavirus 3C protease, Fig. 2). Based on the results of the time course experiments, we hypothesized that the antiviral effect of gemcitabine occurs relatively early in the infection process. To observe which step is influenced by gemcitabine, we performed a time-of-addition experiment. Gemcitabine (10 µM) and rupintrivir (2 µM) were added to the culture medium at −1, 0, 1, 2, 4, 6, 8, 10 and 12 h after virus infection. E30 RNA replication was analyzed at 14 h post-infection.
Treating cells with gemcitabine at -1 h in respect to E30 infection did not inhibit the virus. However, gemcitabine treatment 10 h post-infection effectively suppresses E30 viral infection. In contrast, viral RNA somewhat increased with gemcitabine treatment at 12 h post-infection. This result suggests that gemcitabine did not inhibit E30 replication at an early state of infection, but instead, inhibits the stage of viral infection which occurs at 10 h or earlier post-infection (Fig. 3).
The effects of gemcitabine on the infectivity of E30 particles
The effects of gemcitabine on the infectivity of E30 particles were also investigated. The antiviral activity of E30 pre-incubated (−1 h) with gemcitabine and rupintrivir did not show significant results. In contrast, the sustained presence (0 h) of gemcitabine and rupintrivir during infection resulted in a marked increase in antiviral activity (Fig. 4). This result suggests that gemcitabine does not interact with E30 particles because pre-exposure of the virus to gemcitabine did not alter the infectivity of E30 particles.
NTPs alleviate the antiviral activity of gemcitabine
Previously, it was reported that gemcitabine affects the intracellular levels of nucleotide triphosphate (NTP), especially decreasing CTP levels (22). Thus, we found that the addition of 50 µM ATP, GTP, CTP and UTP significantly reduced the cell viability of RD cells treated with gemcitabine after infection with E30 (Fig. 5A and 5D). We found that the addition of pyrimidine nucleotides, UTP and CTP significantly reduced the antiviral activity of gemcitabine (Fig. 5D). Thus, the addition of gemcitabine limits UTP and CTP levels and may therefore restrict RNA synthesis (Fig. 5C and 5D). Collectively, we found that the anti-viral activity of gemcitabine may be controlled by reducing the levels of CTP and UTP after having been converted into gemcitabine triphosphate.
DISCUSSION
Although vaccines are effective, immunity takes time to develop. Thus, they are of no use to the infected patient (23). Additionally, vaccines must be reformulated each year because of the antigenic drift and rendered ineffective when the formulation does not match the epidemic virus. For these reasons, some antivirals are sought to replace the vaccine by supplying long-continuous protection against both seasonal and pandemic viruses.
Gemcitabine is a nucleoside analog that inhibits cell growth by incorporating into DNA and preventing the action of DNA polymerases. It is approved for the treatment of multiple solid tumors (24). To date, however, the mechanism of action and effectiveness of gemcitabine against E30 have yet to be described. Therefore, we sought to examine whether gemcitabine possesses antiviral activity against E30 using RD cells. Gemcitabine did not show any cytotoxicity at concentrations of 0.4, 2, 10 and 50 µM. Treatment of cells with 10 µM of gemcitabine significantly increased the cell viability to 100%. Therefore, our results support the possibility of gemcitabine as a potential antiviral and therapeutic candidate.
Among the approaches used in determining the antiviral activity of gemcitabine, the time-of-addition assay identified inhibition of viral RNA replication as the mechanism of action. In our study, gemcitabine effectively inhibited the proliferation of echovirus 30, mirroring well-known rupintrivir, an inhibitor of picornavirus 3C protease. Gemcitabine does not suppress the entry of the virus into the cell, but instead inhibits the stage of the viral life cycle that occurs no more than 10h after infection. Therefore, we concluded that the antiviral effect of gemcitabine occurs relatively early during viral infection.
To realize the potential of gemcitabine as an antiviral candidate, its mechanism of activity must be understood. It is important to differentiate between viral inactivation (virucidal activity) from antiviral activity. Direct viral inactivation is an early event where the virus is inactivated before it infects the cells while antiviral activity involves killing the virus or the suppression of viral replication. It would be ideal for viral infection treatments to possess both virucidal and antiviral activities. In our study, gemcitabine did not directly interact with E30 particles as pre-exposure of the virus to gemcitabine did not alter the infectivity of E30 particles. Furthermore, addition of UTP and CTP significantly reduced the antiviral activity of gemcitabine by increasing the levels of pyrimidine nucleotides. The anti-echoviral activity of gemcitabine may be altered by reducing the levels of CTP and UTP after having being converted into gemcitabine triphosphate.
Thus, this study has revealed that gemcitabine could possess antiviral activity against E30. It does not directly inactivate E30 particles, but affects the initial stages of E30 infection by interfering with viral replication. The antiviral activity induced by gemcitabine treatment may be adjusted by reducing the levels of CTP and UTP after it has been converted into gemcitabine triphosphate. These results could be useful in the design of new inhibitors for use as anti-viral agents.



 XML Download
XML Download