

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Fasting and postprandial hyperglycemia of diabetes mellitus can cause diabetic complications such as cardiovascular disease, nephropathy, and neuropathy [1]. Type 2 diabetes, the most common type of diabetes [2], is caused by insulin resistance and relative insulin insufficiency [3]. Type 2 diabetics have 2–10 times greater risk for mortality due to macro- and microvascular diseases compared to non-diabetic population [45]. Because postprandial hyperglycemia is strongly associated with increased risk of cardiovascular complications of type 2 diabetes, it is important to avoid both fasting and postprandial hyperglycemia in type 2 diabetes [6]. Hyperglycemia also increases oxidative stress [7], which is associated with development and progression of cardiovascular diseases related to diabetes mellitus [8].
Postprandial hyperglycemia in diabetes can be controlled by impairing the digestion of dietary carbohydrates by inhibiting α-glucosidases [9]. The α-glucosidase inhibitor acarbose has been used as an oral hypoglycemic agent, but its gastrointestinal side effects include flatulence, abdominal discomfort, bloating, and diarrhea [10]. Therefore, much effort has been directed to the search for safe and effective α-glucosidase inhibitors from natural sources, including edible plants [1112].
Autumn olive (Elaeagnus umbellata Thunb.) is a plant in the family Elaeagnaceae that is native to East Asia [13]. This plant produces small edible deep-red berries with a sweet-tart taste. These are consumed in Korea, China, and Japan [13]. Autumn olive berry (AOB) is good source of vitamin C and vitamin E [1314]. AOB is a rich source of lycopene [15] and has strong antioxidant activity [1416]. Therefore, AOB could be effective in reducing oxidative stress in diabetes mellitus. However, the effects of AOB on α-glucosidase are not clear. AOB is also a good source of proanthocyanidins [13]. The proanthocyanidins in a grape seed extract improve insulin sensitivity by inducing adiponectin mRNA expression in rats with high-fructose diet-induced insulin resistance [17]. The goal of this study was to investigate the anti-diabetic potential of AOB. The α-glucosidase inhibitory activity of AOB was assessed in vitro. The effect of AOB on postprandial glucose excursion was also examined in normal mice. In addition, we studied the effect of chronic treatment with AOB on fasting blood glucose levels, insulin resistance, and adiponectin protein expression in type 2 diabetic mouse model.
Go to : 
MATERIALS AND METHODS
Preparation of AOB extract
Autumn olive berries were purchased from a local market in Hapcheon, Korea. The berries were lyophilized, milled, and then extracted by shaking with ethanol (1:10 m/v) for 12 h twice. After the extract was filtered, ethanol in the extract was removed with a rotary evaporator at 50℃ (EYELA, Tokyo, Japan).
Enzyme inhibition assay
The chromogenic method was used to determine in vitro α-glucosidase inhibitory activity of AOB [18]. The sample solution (5 mg/mL) was prepared by dissolving AOB extract in dimethyl sulfoxide (DMSO; Sigma Chemical, St. Louis, MO, USA). Yeast α-glucosidase (0.7 U, Sigma Chemical) was dissolved in 100 mM phosphate buffer (pH 7.0) which contains bovine serum albumin (2 mg/mL) and NaN3 (0.2 mg/mL) to prepare the enzyme solution. p-Nitrophenyl-α-D-glucopyranoside (p-NGP; 5 mM, Sigma Chemical) was dissolved in 100 mM phosphate buffer (pH 7.0) to prepare the substrate solution. The sample solution (10 µL) and the enzyme solution (50 µL) were mixed and the absorbance was recorded at 405 nm by a microplate reader (MQX200R, BioTek, Winooski, VT, USA). After incubation at room temperature for 5 min, the substrate solution (50 µL) was added to the mixture. The reaction mixture was further incubated for 5 min, and the absorbance was recorded at 405 nm. Acarbose (5 mg/mL, Bayer Korea, Seoul, Korea) was used as a positive control. The assays were done in triplicate.
Measuring postprandial glycemic excursions in mice
A total of 21 male C57BL/6J mice (age, six weeks, Bio Genomics, Seoul, Korea) were separately housed under controlled temperature (19–23℃) and humidity (40–60%) and a 12 h light/12 h dark cycle. The animals had ad libitum access to standard rodent pellets (Purina, Seongnam, Korea) and tap water. Mice weighing 23–26 g were randomly divided into three groups. The control group was administered maltose orally (2 g/kg body weight) after an overnight fast. The AOB group was treated with maltose (2 g/kg) and 500 mg/kg AOB extract. The acarbose group was treated with maltose (2 g/kg) and 50 mg/kg acarbose. Blood samples were acquired from the tip of tail vein 0–120 min after maltose administration. Glucose levels were measured using the Glucotrend glucometer (Roche Diagnostics, Lewes, UK). The areas under the curves (AUCs) for glucose were calculated.
Measuring fasting blood glucose levels in db/db mice
A total of 28 male C57BL/KsJ-db/db mice (age, six weeks) were obtained from the Korea Research Institute of Bioscience and Biotechnology (Ochang, Korea). After 1 week of acclimatization, the mice were assigned to four groups. The control group was fed the basal diet, AIN-93G diet [19] ad libitum for 7 weeks. The low (LAOB) and high (HAOB) autumn olive berry groups received AIN-93G diet containing 0.4% and 0.8% AOB extract, respectively, which replaced cornstarch. Based on a preliminary study, it was assumed that the food intake and average of initial and final body weights of the mice would be 4 g/day and 30 g, respectively. The diets containing 0.4% and 0.8% AOB extract were expected to provide approximately 500 and 1,000 mg/kg body weight of AOB extract daily, based on assumed food intake and average body weight. The acarbose group received the diet containing 0.04% acarbose, which replaced cornstarch. The diet containing 0.04% acarbose was expected to provide approximately 50 mg/kg body weight of acarbose daily. After the mice were fasted overnight (12 h), they were sacrificed by cardiac puncture. Blood samples were collected and serum was obtained after centrifuging the blood at 1,500 g for 15 min. Epididymal white adipose tissue (WAT) was excised. Serum and the WAT were stored at −70℃ for further analyses.
Blood glycated hemoglobin (HbA1C) was measured with a chromatographic method using a commercially available assay kit (BioSystems, Barcelona, Spain). Serum glucose was determined using a commercial enzymatic assay kit (Yeongdong, Seoul, Korea). Serum insulin was assayed using a commercial enzyme-linked immunosorbent assay (ELISA) kit (Mercodia, Uppsala, Sweden). Homeostasis model assessment for insulin resistance (HOMA-IR) was defined as [Fasting insulin (µU/mL) × fasting glucose (mmol/L)/22.5] [20].
To determine the expression of adiponectin using Western blotting, epididymal WAT samples were homogenized in protein extraction solution (PRO-PREP™, Intron Biotechnology, Korea), and then centrifuged at 13,000 g for 30 min. Total protein of the supernatants were measured using a bicinchoninic acid assay (Thermo Fisher Scientific, Rockford, IL, USA) with bovine serum protein as the reference protein. Protein sample (40 µg) was separated using SDS-polyacrylamide gels and then transferred onto a polyvinylidene difluoride membranes (Bio-Rad Laboratories, Richmond, CA, USA) for 30 min. Following blocking using 5% skim milk for 12 h at 4℃, the membranes were incubated with 1:1,000 diluted anti-adiponectin antibody (Cell Signaling Technology, Danvers, MA, USA) and anti-β-actin antibody (Cell Signaling Technology) for 24 h at 4℃. The membranes were incubated with 1:1,000 diluted horseradish peroxide (HRP)-linked anti-rabbit immunoglobulin G (IgG) secondary antibody (Cell Signaling Technology) for 3 h at 4℃. The blots were exposed to enhanced chemiluminescence (ECL) reagent (GE Healthcare, Chalfont, St. Giles, UK). The protein expression was normalized to β-actin. All of the animal protocols used in this study were approved by the Animal Resource Center at Inje University, Korea (Approval n. 2016-020 and 2016-021).
Statistical analysis
The data are presented as the Mean±standard error (SE). One-way analysis of variance with a post hoc Tukey test were used to determine significant differences among the groups (P < 0.05). The statistical analyses were performed with SAS ver. 9.4 (SAS Institute, Cary, NC, USA).
Go to :
RESULTS
In vitro inhibitory effect of AOB on α-glucosidase
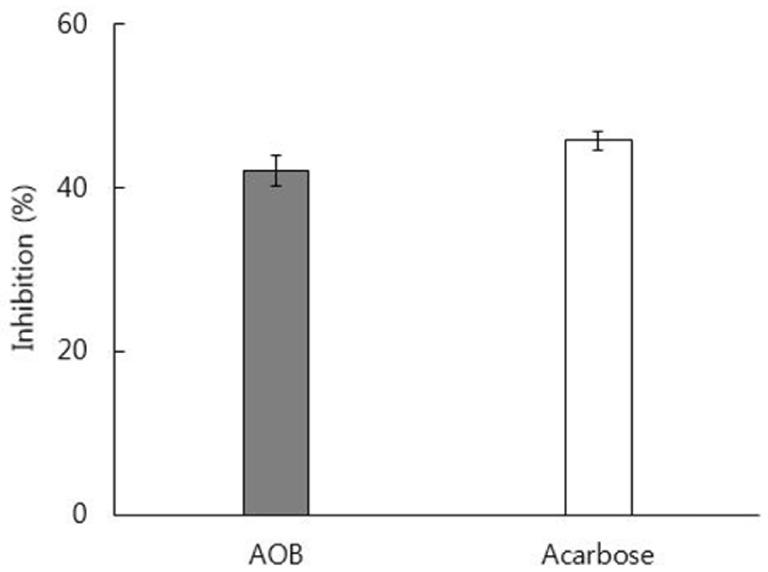
The extraction yield of the ethanol extract of AOB was 36.7%. The AOB extract showed 42.1% inhibitory activity against yeast α-glucosidase, compared to 45.8% inhibition by acarbose, the positive control, in vitro (Fig. 1).
Suppression of postprandial blood glucose in normal mice by AOB
Fig. 2 shows the effects of the AOB extract on postprandial blood glucose levels 0–120 min after maltose loading and the AUCs for the glucose responses in normal mice. The mice treated with the AOB extract at 500 mg/kg showed significantly lowered incremental blood glucose levels 30 (P < 0.01) and 60 (P < 0.05) min after maltose administration (2 g/kg) compared with the mice administered maltose. Administration of acarbose (50 mg/kg) resulted in significant reduction in the incremental blood glucose at 30, 60 (both P < 0.01), and 90 (P < 0.05) min compared with the control group. The incremental blood glucose level at 90 min in mice administered maltose with AOB was not significantly different from those in mice administered maltose alone or maltose with acarbose. The AUCs in the AOB (3,758 ± 482 mg·min/dL, P < 0.05) and acarbose (2,491 ± 221 mg·min/dL, P < 0.01) groups were lower than those in the control group (5,563 ± 620 mg·min/dL). The AUCs of the AOB and acarbose groups did not differ significantly.
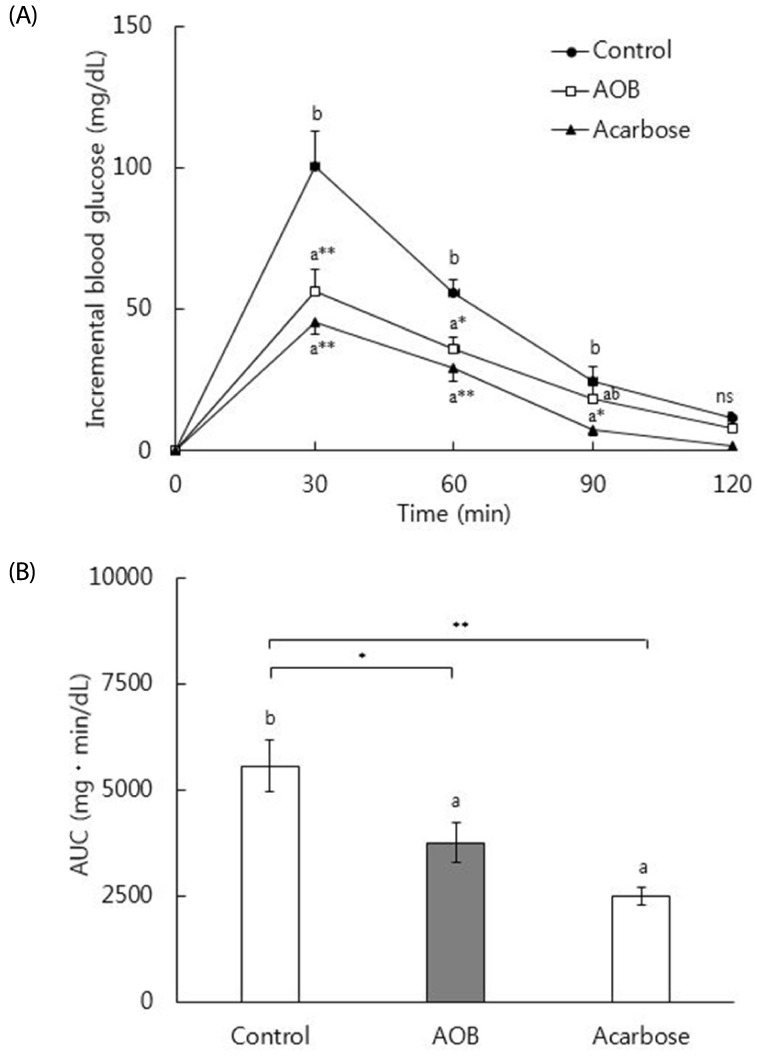 | Fig. 2Increase in blood glucose after administration of extract of autumn olive berry in normal mice.(A) Incremental blood glucose levels. (B) Areas under the glucose response curves. Control group (●): Maltose (2 g/kg) was administered orally to normal mice after an overnight fast. AOB group (□): Maltose (2 g/kg) with extract of autumn olive berry (500 mg/kg) was administered to mice after an overnight fast. Acarbose group (▲): Maltose (2 g/kg) with acarbose (50 mg/kg) was administered to mice after an overnight fast. Values represent Means±SE (n = 7). (A) Means that do not share a common letter are significantly different at P < 0.05 (*) or P < 0.01 (**). (B) Bars that do not share a common letter are significantly different at P < 0.05 (*) or P < 0.01 (**).
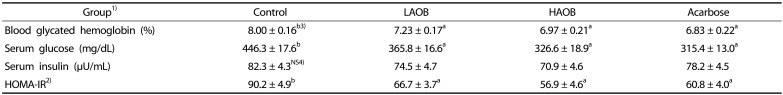
|
Alleviation of fasting hyperglycemia in db/db mice by AOB
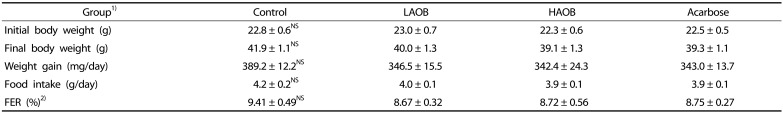
Body weight, weight gain, and feed efficiency ratio were not significantly affected by the consumption of the AOB extract (0.4% and 0.8% wt/wt of diet for LAOB and HAOB groups, respectively) or acarbose (0.04% wt/wt of diet) in the db/db mice (Table 1). The serum levels of glucose were significantly lower in the LAOB, HAOB, and acarbose groups than those in the control group (P < 0.05, Table 2). The consumption of LAOB, HAOB, or acarbose significantly reduced the blood HbA1C compared to the controls. The serum levels of insulin in the four groups did not differ significantly. In comparison with the mice fed the basal diet, the HOMA-IR values of the LAOB, HAOB, and acarbose groups were significantly reduced. Blood HbA1C, glucose, and HOMA-IR values did not differ significantly among the LAOB, HAOB, and acarbose groups. The consumption of HAOB significantly increased the expression of adiponectin protein in epididymal WAT compared to the control and acarbose groups (P < 0.05, Fig. 3). There was a tendency for LAOB to elevate adiponectin expression compared with the control and acarbose groups.
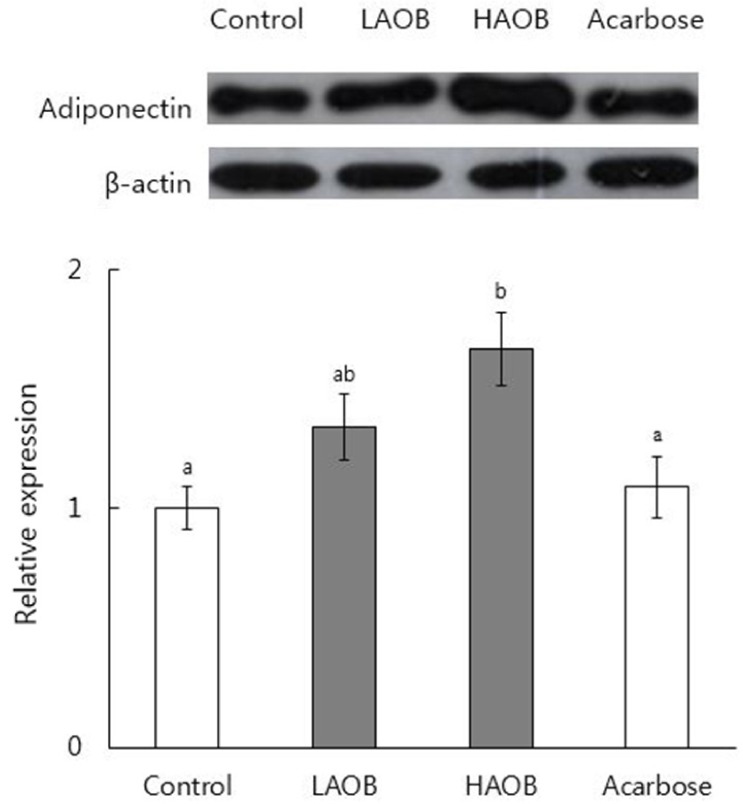 | Fig. 3Expression of adiponectin protein in the epididymal adipose tissue in db/db mice measured by Western blotting.Groups are the same as in Table 1. A representative image of the experiments is shown in the upper panel. The lower panel shows the relative expression normalized to the protein expression levels of β-actin. Values represent Means±SE (n = 7). Bars that do not share a common letter are significantly different at P < 0.05.
|
Table 1
Body weight, food intake, and feed efficiency ratio of db/db mice
1)Control, mice fed AIN-93G diet; LAOB, mice fed diet containing 0.4% autumn olive berry extract; HAOB, mice fed diet containing 0.8% autumn olive berry extract; Acarbose, mice fed diet containing 0.04% acarbose
2)Feed efficiency ratio (FER, %) = (Body weight gain (g/day)/food intake (g/day)) × 100
3)Values are Means±SE (n = 7).
4)NS, not significant
![]()
Table 2
Blood glycated hemoglobin, serum glucose, insulin, and HOMA-IR value of db/db mice
1)Groups are the same as in Table 1.
2)Homeostasis model assessment for insulin resistance (HOMA-IR) = Fasting glucose (mmol/L) × fasting insulin (µU/mL)/22.5
3)Values are Means±SE (n = 7). Means in the same row not sharing a common letter are significantly different at P < 0.05.
4)NS, not significant
![]()
Go to :
DISCUSSION
α-Glucosidase is essential for the digestion of carbohydrates before monosaccharide absorption in the small intestine and inhibition of this enzyme can alleviate postprandial hyperglycemia [9]. The in vitro study demonstrated that α-glucosidase inhibitory activity of AOB extract was 92% as strong as that of acarbose (Fig. 1). α-Glucosidase inhibitors have shown to flatten postprandial increase in blood glucose in both normal mice and mice with diabetes [212223]. In this study we compared the effect of AOB and acarbose on postprandial hyperglycemia after maltose loading in normal mice. The administration of AOB extract at 500 mg/kg was effective at reducing the postprandial blood glucose level and AUC of the glucose response and these effects of AOB were comparable to acarbose administered at 50 mg/kg in the mice (Fig. 2). These results suggest that AOB shows α-glucosidase inhibitory activity in vivo. Postprandial hyperglycemia is seen earlier than fasting hyperglycemia as individuals progress toward type 2 diabetes [24]. Postprandial hyperglycemia is a crucial risk factor for cardiovascular complications among type 2 diabetics [6]. Therefore, flattening the glucose response after a meal with AOB could be beneficial in the prevention of type 2 diabetes and type 2 diabetes-associated cardiovascular complications.
α-Glucosidase inhibitory activity and polyphenolic content of plants are positively correlated [12]. Plant polyphenols, particularly flavonoids, phenolic acids, and tannins, are beneficial in controlling blood glucose by inhibiting α-glucosidase [25]. It has been reported that AOB is high in total phenolic content (190–275 mg/100 g fresh weight) [13]. Polyphenolic compounds in AOB could contribute to α-glucosidase inhibition. It is necessary to find the active components in AOB inhibiting α-glucosidase in further studies.
The effects of different doses of AOB extract on fasting hyperglycemia were examined in db/db mice, which are leptin-deficient and develop obesity, hyperglycemia, and insulin resistance, characteristics of type 2 diabetes [26]. The db/db mice given the AOB extract at 0.4% (LAOB) and 0.8% (HAOB) of the diet for 7 weeks had lower serum levels of glucose in a dose-dependent manner; LAOB and HAOB reduced glucose levels by 18% and 27%, respectively, compared to the control. These hypoglycemic effects of LAOB and HAOB were comparable to those of acarbose given at 0.04% of the diet. The average daily intake of AOB extract by the LAOB and HAOB groups was calculated to be 502 ± 18 and 1,009 ± 19 mg/kg body weight, respectively, whereas the daily intake of acarbose by the acarbose group was 50 ± 2 mg/kg body weight. The mice received LAOB, HAOB, or acarbose showed lowered HOMA-IR, a surrogate parameter of insulin resistance, compared with the basal diet-fed mice. Clinical trials have confirmed that acarbose monotherapy lowers the fasting blood levels of glucose in patients with type 2 diabetes [272829]. Acarbose lowers postprandial hyperglycemia and hyperinsulinemia, thereby reducing the stimulation of insulin synthesis and insulin secretion, contributing to improved insulin sensitivity, which leads to overall glycemic control [30]. The consumption of HAOB increased the expression of adiponectin protein in this study. Adiponectin is a hormone that is expressed in adipose tissues, and adiponectin sensitizes insulin action [31]. Therefore, AOB could improve insulin sensitivity partly by inhibiting α-glucosidase and increasing the expression of adiponectin. HOMA-IR value of HAOB group tended to be decreased compared with the acarbose group, although the difference was not significant. Diabetes increases inflammation which contributes to development and aggravation of diabetic complications [3233]. Adiponectin has been reported to be anti-inflammatory factor which reduces pro-inflammatory cytokine levels [34]. Thus, further study to determine the anti-inflammatory effect of HAOB in diabetes could be worthwhile. The proanthocyanidin-rich extract of grape seed induces adiponectin mRNA expression and the expression of Akt protein, an insulin signaling pathway-related factor, in rats with diet-induced insulin resistance [17]. In our study, the proanthocyanidins in AOB might have helped to alleviate insulin resistance by inducing adiponectin expression.
The consumption of LAOB or HAOB significantly reduced HbA1C. Both fasting and postprandial blood levels of glucose are important for determining HbA1C [35]. HbA1C levels are strongly related to the incidence and severity of diabetic complications, including cardiovascular complications, the most common reason for the premature death of type 2 diabetics [3637]. Consequently, reducing fasting and postprandial hyperglycemia by AOB could be beneficial in prevention of cardiovascular complications.
In conclusion, AOB reduced postprandial hyperglycemia by inhibiting α-glucosidase in normal mice. The chronic consumption of AOB alleviated fasting hyperglycemia and improved insulin sensitivity partly by inhibiting α-glucosidase and upregulating adiponectin expression in db/db mice. Therefore, AOB could be a useful hypoglycemic agent.
Go to :
 XML Download
XML Download