

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Though first described in the early 19th century, classical swine fever (CSF) or hog cholera is a highly contagious pig disease that continues to produce economic losses and agrosecurity threats worldwide. The causative agent, CSF virus (CSFV) is a Pestivirus that belongs to the family Flaviviridae [27]. CSF causes lymphoid depletion by targeting immune cells, which makes the swine more susceptible to other infections [1638]. Clinical signs include high fever, huddling, weakness, drowsiness, anorexia, ataxia (loss of full control of bodily movements), hemorrhage, purple discoloration or cyanosis of the skin, conjunctivitis, and constipation followed by diarrhea [23]. CSF in its most virulent form causes almost 100% morbidity and mortality [40].
Due to the substantial economic impact of a CSF outbreak, research efforts have focused on the development of CSF vaccines that are safe, highly efficacious, fast-acting, cost-efficient, long-lasting, and can differentiate vaccine-induced immunity from field infection. To date, the most frequently used CSF vaccines were derived from the CSFV C strain [17], an attenuated strain from the virulent CSFV Shimen strain [528]. However, modified live virus (MLV) vaccines such as the CSFV C strain vaccine present the challenge of differentiating the MLV-vaccinated pigs from the field-infected pigs. With this constraint, research was directed toward the development of marker CSF vaccines [11]. CSF subunit vaccines are designed to meet the differentiation of infected from vaccinated animals requirement for vaccination [11]. Previous studies have established that the CSFV glycoprotein E2 is the most immunodominant protein in protecting against CSF disease [2225] with multiple identified epitopes [21]. Therefore, E2 subunit vaccines continue to be the focus of CSF subunit vaccine research [67].
Research on CSF E2 subunit vaccines has produced reports of incomplete protection against CSF disease and has shown that inducing E2-specific antibodies is insufficient for conferring protection [39]. This is in contrast to in vitro and in vivo studies that have adequately demonstrated that antibodies against E2 can neutralize and protect against CSF [62526]. Current knowledge of the immune system indicates that there are several, sometimes redundant, pathways by which protective immune responses against a particular pathogen can be induced [12323337]. In subunit vaccine formulations, the immunological component that enhances immune responses, activates multiple immune regulatory pathways, and improves vaccine efficacy are mainly mediated by adjuvants [235]. Hence, adjuvants with the ‘right’ formulation can have the ability to enhance both humoral and cell-mediated immune responses, depending on the mechanism needed to protect against a specific disease [10]. Thus, there is no universal adjuvant that will work in every vaccine. The design or formulation of adjuvants must take into consideration several factors such as antigen, target species, type of immune response, route of inoculation, and safety [18]. Hence, it is possible that formulating the appropriate adjuvant or combination of adjuvants for the immunodominant CSF E2 (antigen) is key to having a highly effective CSF subunit vaccine. We have recently reported on a promising novel E2 CSF subunit vaccine, KNB-E2, which is formulated in an oil-in-water-based adjuvant [29]. KNB-E2 contains insect cell-produced recombinant E2 in a natural conformation, and, with a single dose of KNB E2, vaccinated pigs were clinically protected from CSFV symptoms and they survived virus challenge [29]. KNB-E2-vaccinated pigs can also be easily differentiated from naturally infected pigs by an Erns-specific enzyme-linked immunosorbent assay (ELISA). Upon CSFV challenge, KNB-E2-vaccinated pigs do not produce high levels of Erns antibodies in contrast to the levels in mock-vaccinated pigs [29]. In this paper, we expand on our studies on the CSF subunit vaccine KNB-E2 by determining the effective minimum dose of the E2 antigen, as well as determining the onset and duration of KNB-E2-conferred clinical protection against CSF disease.
Materials and Methods
Animals
Conventional large white-Duroc crossbred weaned specific-pathogen-free male and/or female piglets (3 weeks of age) were purchased from a commercial vendor. Animal care and use protocols were approved by the Institutional Animal Care and Use Committee (IACUC No. 3436) at Kansas State University (KSU). The pigs were fed a standard commercial diet and kept under laboratory biosafety level II conditions at the KSU Large Animal Research Center during the vaccination phase, and under laboratory biosafety level III Agriculture conditions at the KSU Biosecurity Research Institute during the CSF challenge phase. The pigs were randomly allotted into two general groups: KNB-E2-vaccinated groups (n = 5 in each group) and phosphate-buffered saline (PBS; Gibco, USA) vaccinated control groups (n = 5 or 2). The control groups were divided further into PBS-vaccinated, non-challenged (–/–) and PBS-vaccinated, CSFV-challenged (–/+) groups.
CSFV used to infect pigs
The CSF Alfort (HCV Alfort C-718 28) isolate was kindly provided by Dr. Sabrina Swenson from the Animal and Plant Health Inspection Service, United States Department of Agriculture (USDA). The CSFV isolate was passaged seven times in swine testicle (ST) cells (American Type Culture Collection [ATCC], USA) cultured in Dulbecco's modified Eagle's medium (Gibco) supplemented with 10% fetal bovine serum (Atlanta Biologicals, USA) and 1% penicillin-streptomycin solution (Gibco).
Preparation of KNB-E2 and immunization
Expression and purification of recombinant CSFV E2 protein were performed as described previously [29]. The CSFV E2 subunit vaccine, KNB-E2, was prepared by simple hand mixing of purified CSFV E2 with an oil-in-water emulsion adjuvant [15]. This adjuvant containing mineral oil and low-cost plant-based emulsifier is stable for prolonged periods of time at temperatures ranging from 4℃ to 40℃ [15]. Injection sites were monitored and no adverse reactions were observed. One dose of the KNB-E2 prepared for these experiments contained 25 to 75 µg of purified E2 protein per pig, depending on the experiment. All vaccinated pigs were immunized once intramuscularly (neck area) with 2 mL of KNB-E2. The non-vaccinated, non-challenged (–/–) and non-vaccinated, CSFV-challenged (–/+) control group pigs were injected intramuscularly with 2 mL of PBS.
CSFV challenge pig studies
We have previously reported that pigs given one dose of KNB-E2 vaccine containing 75 µg of purified E2 were clinically protected and survived CSFV challenge at 35 days post-vaccination (DPV) [29]. To characterize further the efficacy of the KNB-E2 vaccine, three swine experiments were performed.
Antigen titration (25 µg, 50 µg, and 75 µg of E2 antigen)
Two groups (–/–) and (–/+) of control pigs (n = 5 in each group) were injected intramuscularly with 2 mL PBS and three groups of pigs (n = 5 in each group) were immunized with KNB-E2 formulations (2 mL, intramuscularly) containing 25 µg, 50 µg, or 75 µg of E2 antigen, respectively. Four weeks after vaccination, the pigs were challenged with a 1 × 105 50% tissue culture infective dose (TCID50) of CSFV Alfort (1 mL, intramuscularly).
Onset of immunity (2, 3, and 4 weeks post-vaccination)
Three groups of pigs (n = 5 in each group) were immunized with KNB-E2 vaccine (2 mL, intramuscularly) containing 40 µg of E2 antigen, whereas three groups of (–/+) control pigs (n = 2 in each group) were injected with PBS (2 mL, intramuscularly). Two weeks after vaccination, one group of KNB-E2-vaccinated pigs and one group of (–/+) pigs were challenged with 1 × 105 TCID50 of CSFV Alfort (1 mL, intramuscularly). At 3 weeks post-vaccination, another group of KNB-E2-vaccinated pigs and a (–/+) control group of pigs were challenged with 1 × 105 TCID50 of CSFV Alfort (1 mL, intramuscularly). The third (final) group of KNB-E2-vaccinated and the final (–/+) control group pigs were challenged at 4 weeks post-vaccination with 1 × 105 TCID50 of CSFV Alfort (1 mL, intramuscularly).
Duration of immunity (2 and 4 months post-vaccination)
Two groups of pigs (n = 5 for each group) were immunized with KNB-E2 vaccine (2 mL, intramuscularly) containing 50 µg E2 antigen and two groups of (–/+) control pigs (n = 2 for each group) were injected with PBS (2 mL, intramuscularly). Two months after vaccination, one group of KNB-E2-vaccinated pigs and one group of (–/+) control pigs were challenged with 1 × 105 TCID50 of CSFV Alfort (1 mL, intramuscularly). The second group of KNB E2-vaccinated and the second (–/+) control group pigs were challenged at 4 months post-vaccination with 1 × 105 TCID50 of CSFV Alfort (1 mL, intramuscularly).
Pigs were monitored daily for clinical signs and rectal temperatures after CSFV challenge. Whole blood, serum, and nasal swabs were collected on the day of challenge (0 days post-challenge [DPC]) and every 3 days during the challenge period. All pigs were euthanatized at 14 or 15 DPC.
Anti-E2 specific antibodies were determined by using ELISA as described previously [29]. Briefly, 62.5 ng/mL of purified E2 was used as a coating antigen on 96-well flat-bottomed microtiter plates (Corning, USA). After three washes with PBS containing 0.05% Tween 20 (PBST), diluted sera were added to plates and incubated for 1 h at room temperature. Then, the ELISA plates were washed three times with PBST before detection of the E2-specific antibodies by using horseradish peroxidase-conjugated goat anti-porcine IgG (Southern Biotech, USA). Tetramethylbenzidine (Novex, USA) was used to develop the ELISA plates, followed by the addition of 2 N sulfuric acid (Ricca Chemical Company, USA) to stop the reactions. Optical spectrophotometer readings at 450 nm were measured by using a SpectraMAX microplate reader (Molecular Devices, USA) to determine the relative antibody concentrations in serum samples.
Isolation of RNA and real-time reverse transcriptase polymerase chain reaction for CSFV quantification
Viral RNA from serum samples and nasal swabs were isolated by using a commercial kit (IBI Scientific, USA). Real-time reverse transcriptase polymerase chain reaction (PCR) and analyses were performed by using a StepOnePlus Real-Time PCR System with StepOne software (ver. 2.3; Applied Biosystems, USA) as described previously [1929]. Samples with threshold cycle values equal to or less than 40 were considered positive for CSFV. For virus quantification, passage 7 CSFV Alfort stock (108 TCID50) was serially diluted (108 – 102) before RNA isolation and used to generate a standard
curve.
Serum virus neutralization assay
An indirect fluorescent antibody (IFA) assay was performed to determine the anti-CSFV-neutralizing titers in serum samples of CSFV-challenged pigs. Serum samples were serially diluted (from 5-fold to 10,240-fold) and incubated together with 100 TCID50 of CSFV Alfort for 1 h at 37℃. Residual virus infectivity was determined by adding the serum-virus mixture to 1 × 104 ST cells per well (96-well plate) and incubated for 72 h at 37℃. The cells were then subjected to indirect immunofluorescence staining with CSFV E2-specific murine monoclonal antibody WH303 (APHA Scientific, UK) followed by Alexa-Fluor 488-labeled anti-mouse IgG (Life Technologies, USA). Neutralizing antibody titers in serum samples were expressed as the reciprocal of the highest dilution that causes 50% neutralization.
Blood cell counts
White blood cells (WBC) were enumerated in EDTA-stabilized samples by using a semi-automated animal blood cell counter (VetScan HM5 Hematology system; Abaxis, USA). To ensure instrument quality control, a normal control sample purchased from the supplier was used to calibrate the whole blood analyzer before every use. All samples were analyzed on the day of acquisition.
Results
KNB-E2 vaccine with 25 µg E2 antigen resulted in decreased CSF symptoms upon virus challenge
In order to establish the amount of antigen needed in a single vaccine dose, three groups of pigs were immunized with KNB-E2 containing 25 µg, 50 µg, or 75 µg of CSFV E2 antigen. Pigs were then challenged with 1 × 105 TCID50 of CSFV Alfort at 4 weeks post-vaccination (28 DPV). All KNB-E2-vaccinated pigs (vaccine containing 25 µg, 50 µg or 75 µg of E2 antigen) remained apparently healthy and did not develop fever throughout the challenge phase (panels A and B in Fig. 1). No clinical signs that are typical of CSF infection, such as anorexia (low food intake) or depression were observed in the KNB-E2-vaccinated pigs. In contrast, 3 of the 5 (60.0%) pigs in the (–/+) control group had to be euthanatized before the end of study (on 8 DPC, 10 DPC, and 14 DPC) (panel A in Fig. 1) due to severe clinical CSF symptoms, including high fever (panel B in Fig. 1), loss of body weight (data not shown), seizures, diarrhea, and severe leukopenia (panel C in Fig. 1). All (–/+) control pigs became viremic after challenge, exhibiting up to 107 TCID50 CSFV detected in blood (panel A in Fig. 2) and nasal cavity (panel B in Fig. 2). In contrast, a few of the KNB-E2-vaccinated pigs, notably even some in the 25 µg group, had low transient levels of CSFV, which were eventually cleared by the KNB-E2-vaccinated pigs within the study duration (panels A and B in Fig. 2).
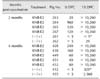
Comparable levels of CSFV E2-specific antibodies were detected in the serum (dilution 1/1,000) by ELISA in all three KNB-E2-vaccinated groups within 14 DPV (panel C in Fig. 2). However, a lower level of E2-specific antibodies was observed in pigs vaccinated with 25 µg of E2 antigen at 21 DPV (panel C in Fig. 2). After CSFV challenge, all three KNB-E2-vaccinated groups displayed an anamnestic response characterized by a rapid increase of E2-specific antibodies in the serum (dilution 1/10,000; panel D in Fig. 2) starting at 6 DPC. Similarly, the serum anti-CSFV-neutralizing antibody titers (VNA) detected by IFA in pigs vaccinated with 25 µg of E2 antigen were slightly lower at 28 DPV/0 DPC (i.e., before challenge), compared to that in pigs vaccinated with 50 µg or 75 µg of E2 antigen (Table 1). However, by the end of the study period (15 DPC) all KNB-E2 vaccinated pigs developed very high (≥ 10,240) VNA titers. These data indicate that the levels of E2 antibodies and VNA titers in KNB-E2 pigs vaccinated with 25 µg of E2 antigen were sufficient for clinical protection against CSF disease.
In addition, 2 of the 5 pigs in the (–/+) control group (pigs 168 and 170) survived the CSFV challenge. These two pigs had > 40℃ fever at 3 DPC to 6 DPC along with loss of appetite and depression. The viremia detected in pigs 168 and 170 peaked at 6 DPC and was notably lower at 9 DPC; however, CSFV was still detected in these pigs at the end of the study (panels A and B in Fig. 2). Pigs 168 and 170 had no detectable VNA titers before challenge (0 DPC; Table 1); hence, there was no pre-existing immunity against CSFV. Interestingly, these (–/+) control pigs both developed low detectable VNA titers at 15 DPC but to a much lower degree than that in the KNB-E2-vaccinated pigs (Table 1). Collectively, our data suggest that KNB E2 vaccine containing as little as 25 µg of E2 antigen can confer clinical protection against CSFV infection in swine.
KNB-E2 immunized pigs were clinically protected from CSF as early as 2 weeks post-vaccination
Vaccination was carried out in three groups of pigs with the KNB-E2 formulation containing 40 µg of CSFV E2 antigen. The first group of KNB-E2-vaccinated pigs was challenged with CSFV at 2 weeks post-vaccination. The remaining groups of KNB-E2-vaccinated pigs were challenged at 3 or 4 weeks post-vaccination. Age-matched (–/+) PBS-vaccinated pigs were also challenged at 2 weeks, 3 weeks, and 4 weeks post-vaccination as control groups. All KNB-E2-vaccinated pigs at 2 weeks, 3 weeks, and 4 weeks post-vaccination remained apparently healthy after they were challenged (panel A in Fig. 3). The 2-week KNB-E2-vaccinated group developed fever at 3 DPC similar to that in the (–/+) control group (panel B in Fig. 3). The 3-week and 4-week KNB-E2-vaccinated pig groups developed transient low fevers at 5 DPC. However, from 8 DPC onwards, all KNB E2-challenged pigs recovered and had slight or no fever (panel B in Fig. 3); moreover, they did not further develop other clinical signs of CSF. In contrast, the (–/+) challenged control animals developed typical clinical signs of CSF; and 5 of 6 (83.3%) of the (–/+) control pigs had to be euthanatized between 6 DPC to 8 DPC (panel A in Fig. 3).
All of the KNB-E2-vaccinated, CSFV E2-challenged pigs (2-week, 3-week, and 4-week groups) steadily gained weight throughout the 14-day challenge period with only a minimal decrease in weight gain observed in the 2 week pig group between 3 DPC and 6 DPC (panel C in Fig. 3). In contrast, weight loss was observed in the (–/+) challenged control group starting at 3 DPC (panel C in Fig. 3). At 8 DPC, only one (–/+) control pig remained (pig 313) and this pig appeared to have recovered from CSF (panels B and C in Fig. 3). Since all pigs were CSF-free, the steady weight gain of pig 313 toward the end of the study suggested the presence of an innate resistance or a conversion into a subacute or chronic disease. Similarly, a decrease in leukopenia was observed in pig 313 starting at 9 DPC (panel D in Fig. 3) and continuing until the end of study period, suggesting recovery from CSFV challenge. On the other hand, slight decreases in WBC numbers were observed in all KNB-E2-vaccinated pigs (2-week, 3-week, and 4-week groups) at 3 DPC and 6 DPC; but these numbers were recovered by 9 DPC (panel D in Fig. 3).
CSFV RNA was detected early in the 2-week KNB-E2-vaccinated pigs but viremia cleared by 12 DPC (panel A in Fig. 4). During the challenge period, CSFV RNA was detected in 2 of 5 pigs in the 2-week KNB-E2-vaccinated group; 1 out of 5 in the 3-week KNB-E2-vaccinated group; and none in the 4-week KNB-E2-vaccinated group at the end of the study period (panel A in Fig. 4). On the other hand, CSFV RNA was not detected in the nasal cavities of any of the KNB-E2-vaccinated pigs (2-week, 3-week, and 4-week groups) at 9 DPC (panel B in Fig. 4). At 9 DPC, in which only pig 313 remained, CSFV RNA was still detected in this pig. Pig 313 remained CSFV RNA positive throughout the challenge period. However, this pig appeared to be recovering from CSF by the end of the study (panel B in Fig. 4).
The 2-week KNB-E2-vaccinated pig group demonstrated detectable but lower levels of E2-specific antibodies in serum (dilution 1/10,000) compared to the levels in the 3-week and 4-week KNB-E2-vaccinated groups (panel C in Fig. 4); interestingly, no other CSF clinical signs were observed in the 2-week KNB-E2-vaccinated CSFV-challenged pigs. Likewise, the 2-week KNB-E2-vaccinated group had no or little VNA titers on the day of challenge, but the pigs developed sufficient protective titers of anti-CSFV-neutralizing antibodies by the end of the experimental period (Table 2). Collectively, our results suggest that even though some vaccinated pigs were positive for CSFV RNA and had transient fever, the KNB-E2 vaccine conferred clinical protection against CSF disease at 2 weeks post-vaccination.
KNB-E2-immunized pigs were clinically protected from CSF disease at 2 months and 4 months post-vaccination
The duration of KNB-E2 vaccine efficacy was determined by challenging KNB-E2-vaccinated pigs with CSFV at 2 months and 4 months post-vaccination. In this pig study, one pig out of the 5 pigs in the 2-month KNB-E2 group had to be euthanatized during the vaccination phase due to a health problem unrelated to CSF. As with the previous pig experiments, at 2 months and 4 months post-vaccination, all KNB-E2-vaccinated pigs survived CSFV challenge (panel A in Fig. 5). In contrast, only 50% of the (–/+) PBS-vaccinated control pigs survived the CSFV challenge (panel A in Fig. 5). The 2-month and 4-month KNB-E2-vaccinated pig groups developed transient one-day fever at 5 DPC and 6 DPC, respectively (panel B in Fig. 5). The challenged (–/+) control pigs had elevated temperatures of up to 41.5℃. Interestingly, the (–/+) control pigs that survived the CSFV challenge exhibited higher fevers than those of the (–/+) control pigs that had to be euthanatized (data not shown).
Continued body weight gain was observed in both the 2-month and 4-month KNB-E2-vaccinated pigs (panel C in Fig. 5) after CSFV challenge. At 10 DPC, only two of the (–/+) control pigs remained, and these pigs seemed to have recovered as evidenced by their body weight gain observed starting at 9 DPC and continuing to the end of the study period (panel C in Fig. 5). Interestingly, the 4-month KNB-E2-vaccinated pigs exhibited higher WBC counts than 2-month KNB E2-vaccinated pigs at 0 DPC (panel D in Fig. 5). A slight decrease in WBC numbers was observed in all 2-month and 4-month KNB-E2-vaccinated pigs immediately after CSFV challenge but these numbers were recovered after 6 DPC (panel D in Fig. 5).
Both the 2-month and 4-month KNB-E2-vaccinated groups had CSFV-positive serum at 6 DPC but both groups cleared the CSFV at 9 DPC (panel A in Fig. 6). In the nasal swab samples, the 4-month KNB-E2-vaccinated group had a higher level of CSFV detected than that in the 2-month KNB-E2 group but was clear of the virus at 12 DPC (panel B in Fig. 6). One pig in the 2-month KNB-E2-vaccinated group and two pigs in the 4-month KNB-E2-vaccinated group did not develop indications of viremia throughout the challenge phase (data not shown). Together with the observed higher WBC counts in the 4-month KNB-E2-vaccinated pigs (panel D in Fig. 5), the results suggest an age-related factor in the immune responses against CSFV.
At 0 DPC, the 4-month KNB-E2-vaccinated pigs demonstrated lower levels of E2-specific antibodies than that in the 2-month KNB-E2-vaccinated pigs and a level comparable to that in the (–/+) control pigs from 0 DPC to 6 DPC (panel C in Fig. 6). Interestingly, by 9 DPC, the 4-month KNB-E2-vaccinated group produced higher amounts of E2-specific antibodies than the 2-month KNB-E2-vaccinated group (panel C in Fig. 6). Similarly, the 4-month KNB-E2-vaccinated pigs demonstrated lower levels of VNA titers at 0 DPC than those in the 2-month KNB-E2-vaccinated pigs (Table 3). However, by the end of the challenge period (15 DPC), both 2-month and 4-month KNBE2-vaccinated pig groups developed very high titers (≥ 10,240) of VNA. Taken together, our data suggest that KNB-E2-immunized pigs are clinically protected against CSF disease for
at least 4 months post-vaccination.
The 2-month (–/+) PBS-vaccinated control pig (pig 262) that survived the CSFV challenge developed low VNA titers (Table 3). On the other hand, the older two pigs in the 4-month (–/+) group (pig 432 euthanatized at 10 DPC and pig 455 that survived the CSFV challenge) both exhibited detectable levels of VNA (Table 3) after CSFV challenge. Unlike younger pigs, the 4-month (–/+) control pigs started to develop VNA within 6 DPC and 9 DPC (< 5 anti-CSFV neutralizing bodies that neutralized 50% [ND50] of virus at 6 DPC and 30 ND50 at 9 DPC). In contrast, no E2-specific antibodies were detected by ELISA in these two 4-month (–/+) control group pigs throughout the study period (panel C in Fig. 6). These results suggest an age-related factor that may have an effect on CSF survival.
Discussion
We have demonstrated that KNB-E2, composed of a
recombinant E2 protein expressed by insect cells and an oil-in-water emulsion-based adjuvant, is safe to administer and can effectively stimulate protective immunity against CSF with a single dose. Traditional subunit vaccines generally require multiple doses to be effective. For CSF, only two licensed subunit vaccines are commercially available and only in a few countries (Porcilis pesti [MSD Animal Health, USA] and Advasure, formerly Bayovac CSF E2 [Pfizer, USA]) [45]. These CSF subunit marker vaccines effectively induce CSF protection [1], but two doses are required followed by single-dose re-vaccination every six months. The efficacy displayed by the KNB-E2 vaccine illustrates the importance of having a potent adjuvant and an optimal interaction between vaccine components [14]. In the present study, antigen titration of the recombinant CSFV E2 glycoprotein showed the development of high CSFV-neutralizing titers and protection from CSF clinical symptoms with a single KNB-E2 containing 25 µg of E2. This antigen amount is less than the minimum 32 µg of E2 glycoprotein per dose in Advasure. During the vaccination phase, the 25 µg KNB-E2-vaccinated pigs had lower E2-specific antibodies and VNA compared to those in pigs vaccinated with 50 µg and 75 µg of KNB-E2. However, the level of protection conferred by the 25 µg KNB-E2 was sufficient to clinically protect against CSF disease as demonstrated by the absence of typical CSF pathology and the high E2-specific antibody neutralizing titers detected at the end of study period.
The onset of protection of the KNB-E2 vaccine was determined to be at approximately 2 weeks post-vaccination, based on the protection against mortality and the reduction of CSF clinical signs. Although the 2-week KNB-E2-vaccinated pigs had very low antibody titers at the time of challenge and showed mild signs of the disease (transient elevated temperatures), these pigs had developed sufficient immunity to adequately respond to a CSFV challenge. The dynamics of the anti-CSFV-neutralizing antibody produced within 2 weeks post-vaccination is consistent with results that were previously reported [631]. The differential efficacy of KNB-E2 at 2 weeks, 3 weeks, and 4 weeks post-vaccination may be due to the insufficient maturation of the humoral immune response in young pigs. The cellular immune responses that play an important role to control CSFV [1739] were not addressed in this paper; however, evidence suggests that, with optimal antigen and adjuvant formulations, subunit vaccines can induce both humoral and cellular-mediated immune responses [1035]. A representative oil-in-water adjuvant is MF59 from Novartis (Switzerland). Its proposed mechanisms for immune enhancement include efficient antigen delivery, immune potentiation at the injection site resulting in early leukocyte recruitment, and activation and maturation of antigen-presenting cells that result in antigen presentation through MHC molecules that subsequently allow T-cell recognition and B-cell activation. In addition, MF59 is reported to be more potent than aluminum-based adjuvants in terms of both antibody and T-cell responses [34]. Future studies will be conducted to investigate the role of cellular immunity in KNB E2 mediated protection against CSFV.
An important marker of an effective CSF vaccine is the period during which pigs are protected after vaccination. Earlier investigations demonstrated that CSF subunit vaccines can induce protective immunity that lasts for up to several months [820]. The results presented in our study clearly demonstrate that KNB-E2 can confer protection against CSFV at least 4 months post-vaccination. With the high levels of E2-specific and CSFV-neutralizing antibodies induced, we speculate that pigs vaccinated with KNB-E2 would be clinically protected against the disease for more than 4 months. Even though a low VNA was detected in the 4-month KNB-E2-vaccinated pigs before challenge, interestingly, we observed higher anti-CSFV-neutralizing antibodies in the 4-month KNB-E2 pigs than that in the 2-month KNB-E2 pigs after CSFV challenge. It appears that the CSFV challenge induced an anamnestic antibody response and, therefore, provided clinical protection against CSF disease.
Of interest in this study are the PBS-vaccinated, CSF-free pigs that survived the CSFV challenge. The apparent development of low VNA titers in these pigs toward the end of the challenge study, suggests that the innate immune system may have a critical role in the CSFV challenge. Similar to humans, pigs have been reported to exhibit substantial genetic variation in immunity [1330]. Hence, the differences in anti-CSFV immunity observed in the present study may be attributed to genetic diversity in the selected pigs.
On the other hand, age-dependent variation in porcine immune responses to viral infection has been reported [2436]. Therefore, the restriction of CSFV growth observed in the pigs challenged at 4 months post-vaccination may be attributed to an age-related innate immune resistance. Previous studies have reported stronger cell-mediated responses in older pigs; in contrast, induced humoral responses were unrelated to animal age [24]. In this study, we observed that older pigs may have the capacity to quickly mount an anti-CSFV-specific VNA response starting at approximately 6 DPC. Since no E2-specific antibodies were detected in the older pigs throughout the study, these results suggest that anti-CSFV protection could be conferred by the more potent innate immune mechanisms in mature pigs.
Protection against transmission was not investigated in this study, but single-dose KNB-E2-vaccinated pigs did transiently test positive for CSFV RNA. Hence, there remains a certain risk of vertical transmission. Nevertheless, our results suggest that KNB-E2 reduces clinical CSF signs and might be valuable in reducing CSF related losses. Another advantage of using this vaccine is the capacity to discriminate between vaccinated and CSF-infected pigs [29]. However, to ensure complete protection against CSF, a second KNB-E2 dose might be beneficial. An improved, more potent adjuvant or incorporation of multiple immunogenic antigens could also be considered.
Our results have demonstrated that CSF subunit vaccine KNB-E2-mediated protection is dose dependent. A minimal amount of E2 antigen is required for this subunit vaccine to confer rapid and long-lasting clinical protection against CSF. Further evaluation of the efficacy of KNB-E2 against current strains of CSFV will shed light on the potential role of KNB-E2 in reducing CSF related losses.








 XML Download
XML Download