

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Olfaction is essential for survival and enhances quality of life as it is indispensable for the acquisition of various emotions, episodic memories, and social behaviors [3156]. A variety of specialized neurons responsible for olfactory information processing are localized in the olfactory epithelium (OE), the olfactory bulb (OB), and in other higher order olfactory centers such as the piriform cortex and the orbitofrontal cortex [68]. Anatomical or molecular disturbances in this system can lead to olfactory impairment, either as conductive or sensorineural loss. Conductive impairment is caused by mechanical obstruction of the olfactory apparatus preventing the delivery of odor stimulant to the OE, while the latter is associated with abnormal processing of stimuli in the OE and central olfactory system due to a defect in the neuronal network [2446]. Olfactory dysfunction is a very common clinical condition with a high prevalence; Lee et al. [66] reported that 4.5% of adults (range, 20–95 years) have self-awareness of olfactory dysfunction in Korea. The clinical implications of olfaction have been often unnoticed or underestimated, and recovery from olfactory loss remains challenging with limited therapeutic options, especially in the case of sensorineural olfactory defects [87].
Although sinonasal disease and upper respiratory infection, as well as head trauma, are the most commonly reported clinical manifestations leading to transient or persistent olfactory disturbance [244656], the aging process has been regarded as a major contributing factor for anosmia in general [86186]. It has been reported that more than half of ‘healthy’ individuals over age 65 have a decline in olfaction, and the number of victims increases steeply with aging to nearly 80% in those over 80 years of age [41]. In addition, patients with a variety of neurological diseases including Alzheimer's disease (AD), Huntington's disease (HD), Parkinson's disease (PD), multiple sclerosis, and dementia show a high susceptibility to olfactory dysfunction [948]. Interestingly, olfactory loss seems to precede the onset of other typical neurodegenerative symptoms such as memory loss and motor impairment, indicating the importance of olfactory assessment for an early diagnosis of various neurological disorders [7885101]. Therefore, olfactory disorders have become a matter of societal as well as individual importance in an era of accelerated population aging with concurrent increasing risks of neuropathological medical conditions [1834]. Moreover, elucidating the underlying mechanisms of olfactory loss might be necessary not only to understand the nature of neurodegeneration but also to develop disease management strategies for treatment and prevention.
Age-related physiological changes such as an increased susceptibility to infection, long-term exposure of air-borne agents (e.g., smoking, air pollutant), and blood flow reduction in the nasal mucosa can cause direct damage to olfactory neurons [4088]. Autopsy and brain imaging have revealed gradual atrophy of the OB accompanied by the loss of olfactory nerve fibers with aging [1725]. Similarly, functional MRI and electroencephalography recordings have demonstrated a neurodegeneration-associated decline in olfactory responsiveness with decreased neural activity in patients [1327286999]. Some researchers have suggested a correlation between the development of typical neuropathological signs such as protein aggregates and the severity of olfactory dysfunction; post-mortem brain studies have shown that amyloid beta plaque and neurofibrillary tangles were often accumulated in the OB neuronal dendrites in AD patients [57102], and intracytoplasmic Lewy bodies induced by α-synuclein aggregation were identified in the OE and OB neurons of PD patients [2135], while others found no significant correlations [10]. In addition, defects in neurotransmitter signaling seem to impede proper neural interactions for olfactory processing in a context-dependent manner. In particular, acetylcholine might play a critical role in maintaining normal olfaction in humans [1419]; indeed, Bohnen et al. [19] reported that enhanced cholinesterase activity not only decreased acetylcholine concentration but also led to olfactory decline. On the other hand, donepezil, a cholinesterase inhibitor, can restore the impaired olfaction of AD patients [12,97]. Interestingly, despite dopamine deficiency being regarded as a key pathophysiologic feature of PD, neither dopaminergic neuronal denervation in the OB nor treatment with dopaminergic agents has improved olfaction of PD patients, indicating the complicated nature of region-specific neurodegenerative processes [319]. Therefore, assessment of the brain via indirect imaging or post-mortem studies would be insufficient in explaining in detail how olfactory dysfunction begins and develops during aging and neurodegeneration. In this regard, rodent models have significantly contributed to the establishment and evaluation of hypotheses in these fields.
Animal Models for the Study of Olfactory Impairment
Animal models are indispensable in the investigation of disease pathogenesis and the development of relevant therapeutics. Rodent models are the most preferred experimental species due to their genomic similarity to humans, convenience for genetic modification, and cost-effectiveness. Rodent models can recapitulate the neurological molecular pathology including the production of protein aggregates and alterations in the neurotransmitter system. Moreover, they have been widely used to distinguish between cause and effect and to determine the clinical significance of novel findings by modulating (either up- or down-regulation) specific signalings in vivo.
Olfaction is gradually impaired with age or neurodegeneration in rodents, and, in both rodent and human, the cellular mechanisms of olfactory dysfunction have been studied. It has been reported that olfactory loss during aging is associated with a reduction in the number and diversity of olfactory neurons caused by: i) cell cycle arrest in the OE neuroblast; ii) significant decline in neurogenesis in the subventricular zone (SVZ); or iii) transcriptional down-regulation in olfactory receptor subtypes [657089]. In neurological disease models, toxic metabolites and protein aggregates with secondary neuroinflammatory molecules seem to accelerate olfactory deficit (details presented in the following). Therefore, rodent models can provide valuable information regarding cellular and molecular mechanisms associated with olfactory impairment for application in establishing therapeutic strategies.
Methods for olfactory evaluation in mice models
To date, various psychophysical, electrophysiological, and psychophysiological tests for olfactory function have been developed to quantify and qualify olfaction in the clinic [1339]. Because of its relatively simple and intuitive procedure, psychophysical olfactory testing has been widely used, with some modifications, to evaluate the olfaction of experimental animals. Odor recognition, identification, and discrimination abilities are the key criteria for olfactory assessment using psychophysical olfactory tests.
In this review, we briefly introduce the characteristics and principle procedures of the most frequently used olfactory tests for experimental mice (for more detail, refer to [1538103107]). Note that in these tests, each subject should perform the task once to avoid errors derived from unintended behavioral learning rather than assessing pure olfaction, except in the case of testing long-term olfactory memory formation ability.
Olfactory sensitivity test (olfactory threshold test, olfactory preference test, and olfactory avoidance test
In these tests, animals are simultaneously exposed to unfamiliar, novel odor cues and control odor cues such as water. During testing, various concentrations of the odors are introduced to subjects in a descending order (from high to low concentration) and the total exploring times between stimulus and water cues are compared to determine ‘olfactory sensitivity’. In general, subjects with normal olfactory function are expected to spend more time investigating the higher stimulus concentration than the lower concentration of the stimulus or the water control.
The basic test format can be modified for use in advanced testing such as testing olfactory preference and olfactory avoidance. These tests are aimed at assessing the ability to distinguish between attractive (e.g., vanilla or peanut butter scent are preferred) or aversive scents (e.g., 2-methylbutyric acid) with the results based on total exploratory times.
Olfactory discrimination test (olfactory habituation-dishabituation test [non-social recognition test], social recognition test [block test], and olfactory memory test)
These tests are useful in determining whether mice can discriminate a novel odor stimulus from familiar ones. During the olfactory habituation-dishabituation test, which is considered a basic olfactory discrimination test, subjects are first presented with odor ‘A’ for habituation. Subject response to odor A decreases with repeated exposure to the odor (i.e., habituated behavior). When a different odor, ‘B’, is introduced to mice during this test procedure, exploring time to the novel stimulus will be increased (i.e., dishabituated behavior).
With some modification, this test can be applied as a ‘social recognition test’ to determine whether mice can discriminate ‘self’ and ‘non-self’ by using olfaction (note that the original olfactory discrimination test is also known as ‘non-social recognition test’ in contrast to the social recognition test). In the social recognition test, the subject is exposed to blocks from its own cage and from another non-self cage and exploration time for each block is quantified. A standard wild-type mouse always tends to show a preferential interest in the block from the non-self, foreign animal.
Finally, habituation-dishabituation behavior is also used for the assessment of olfactory memory by evaluating the ‘rewarded olfactory discrimination’ ability of subjects, although this test is usually used to evaluate cognitive function rather than olfaction itself.
Olfactory perception test (buried food test)
The buried food test is one of the most frequently used tests for olfactory evaluation and is useful in determining an animal's olfactory perceptibility. Prior to the test, animals undergo food restriction to increase food motivation behavior. Then the subject is introduced into the test cage with food buried under the bedding, and the latent period to locate and eat the food is recorded. Control testing with ‘exposed’ food (i.e., located on the bedding) should be performed in parallel with the buried food test to confirm that subject is free from any appetite disorder or visual defect. Typically, mice with olfactory deficit show an increased latency in the buried food test but not in the exposed food test.
Contribution of Microgliosis to Olfaction
Microglial function in healthy and pathogenic brains
The central nervous system (CNS) consists of neurons and supportive glial cells including oligodendrocytes, astrocytes, and microglia. Unlike other CNS cells that are derived from an ectodermal lineage, microglia are known to originate from the mesodermal hematopoietic lineage, and their primitive progenitors in the yolk sac migrate into the developing brain during early embryogenesis [47]. Microglia perform a constant surveillance activity, thereby being considered as the resident macrophages in the CNS, and they maintain their population via consistent local proliferation throughout the lifetime of the CNS [1].
In the normal mature CNS, the majority of microglia are under a steady state with ramified morphology; so-called ‘resting microglia.’ In addition to their immune surveillance role, resting microglia contribute to many biological processes in the CNS including neurogenesis [93]. It has been previously reported that microglia participate in the arrangement of appropriate neural circuits by removing cell debris during the embryonic period and early postnatal life [6], and, even in the resting state, microglia residing near a neurogenic center such as the OB and subgranular zone are reported to perform phagocytosis of apoptotic cells during neurogenesis [8493]. When microglia are activated in response to various environmental stimuli, they expand their population (microgliosis) as well as transform into an amoeboid-like morphology with increased secretion of cytokines and inflammatory mediators. Indeed, almost every neuropathic condition is accompanied by extensive microglial activation and profound microgliosis [5775106]. Activatation of microglia is often divided into two states, classical activation (M1) and alternative activation (M2), based on the activating factors (M1: lipopolysaccharide [LPS] and interferon γ [IFN-γ]; M2: interleukin [IL]-4 and IL-13), expressed cell specific markers (M1: CD86; M2: CD206), and the main biological functions (M1: production of proinflammatory molecules; M2: enhanced phagocytosis); however, there are some limitations in this M1-M2 microglial classification system that mainly stem from the fact that the system overlooks the plasticity and complexity of microglia and because there is no actual boundary between the two phenotypes [71]. Microglia can provide both beneficial and detrimental effects on neurodegenerative process depending on the context; however, it is generally accepted that M1 microglia aggravate an on-going neuropathology by producing neurotoxic factors, while M2 microglia help to resolve the problem via phagocytosis of unwanted molecules. For instance, classically activated microglia-derived proinflammatory cytokines such as IL-1β and tumor necrosis factor α (TNF-α) reduce the number of newborn cells in neurogenic centers and impede normal synaptic activity in neurons, as is frequently observed in aging and neurodegenerative disorders [76]. High levels of nitric oxide and reactive oxygen species also contribute to microglia-induced neural apoptosis [3367]. The M1 microglia-derived proinflammatory signals also stimulate neighboring astrocytes to proliferate and produce cytotoxic factors, resulting in more severe neuroinflammation [98]. In contrast, several studies have shown that phagocytotic ability of alternatively activated microglia is important to prevent/delay the development of neurodegeneration by removing the causative agents in several neurologic disorders (e.g., amyloid beta in AD) and restricting the affected lesions [5860]. Therefore, comprehensive understanding of microglial roles in various neuropathologic conditions should precede testing of the therapeutic potential of candidate drugs that modify microglial activity.
Microglial activation in the olfactory system
One of the main neurogenic centers, the SVZ, possesses adult neural stem cells and produces neural progenitors upon stimulation. These neuroblasts traverse the rostral migratory stream to the OB, where they mature and differentiate into granule cells and periglomerular neurons [22]. Although there is debate on the presence of OB neurogenesis in human adults, it has been reported that newly generated OB neurons seem to be essential for the maintenance of normal olfaction in animals because a chronic treatment of the antimitotic agent AraC ablated SVZ-derived neurogenesis and was followed by olfactory deficits in mice [22].
It has been previously reported that microglia participate in the arrangement of the appropriate neural circuits during neurogenesis by removing cell debris during the embryonic period and postnatal life [684]. As part of the SVZ neurogenic center, therefore, the OB contains a high number of microglia, which allows investigation of the role of microglia in neural differentiation, survival, and apoptosis as well as microglia-neuronal interaction [80]. The number of phagocytic amoeboid-like microglia in the rat OB gradually increases during the first 3 to 4 weeks after birth then decreases as structural and functional stabilization of OB circuitry becomes established [2642]. Recently, Ribeiro Xavier et al. [77] reported a distinctive subtype of microglia within the SVZ-OB axis. These microglia had an amoeboid-like morphology with an enlarged cell body and produced M2-like cytokines such as IL-4 and IL-10. It has also been noted that microglial depletion via intravenous injection of Cd11b-conjugated saporin toxin resulted in a decreased number of OB neuroblasts migrating from the SVZ in rodents, indicating that appropriate microglial activation might be essential for the survival and migration of neuroblasts in the SVZ-OB axis [77].
In addition, OB neural damage is often accompanied by neuroinflammation in response to pathologic stimuli. In general, microgliosis and astrocytosis develop simultaneously during the inflammatory process. Some studies have shown the morphological diversity of astrocyte subtypes and their expression of synaptogenetic molecules in the OB layers, implying that astrocytes can also contribute to OB structure formation during the developmental stage [111245]; however, direct evidence of astrocytic involvement in olfactory loss and their possible roles in OB pathogenesis has not been reported yet. On the other hand, the correlation between microglial activity and the development of olfactory dysfunction in various pathologic conditions has been discussed, as summarized in the following.
Olfactory deprivation models (naris occlusion model and chemically-induced OB ablation model)
It has been reported that occluding one naris during the early postnatal period significantly alters olfactory neuronal connectivity, leading to an olfactory deficit [23]. Histologically, unilateral naris closure has resulted in a decline in OB size due to massive loss of tyrosine hydroxylase-positive dopaminergic interneurons in the OB [50]. Notably, progressive microgliosis within the OB was also observed with the morphological transformation from resting state into the amoeboid state, indicating increased phagocytic activity. As a result, most adult-born OB neurons are engulfed by microglia in the occluded OB, leading to the loss of synaptic activity and olfactory dysfunction [3650].
Dichlobenil administration is another widely used method for OB deafferentation as it leads to a rapid degeneration of olfactory sensory neurons (OSNs) by disrupting their axonal projections [63105]. Importantly, it is reported that dichlobenil-induced ablation of OE neurons not only decreased the survival of newly generated neurons in the OB but also increased the proliferation of microglia (but not of astrocytes) similar to the results obtained from a unilateral naris closure model [63]. To determine the cause and effect between olfactory loss and microgliosis, the anti-inflammatory agent minocycline was administrated to dichlobenil-treated mice, then, olfactory behavioral tests were conducted. It was reported that minocycline treatment could prevent neuronal apoptosis and olfactory impairment, indicating that suppression of microglial activity is protective of olfactory deafferentation and functional deficits after olfactory ablation [63].
Neuroinflammation models (inducible olfactory inflammation and bacteria-infected model)
A prolonged inflammatory status such as chronic rhinosinusitis is a common predisposition to olfactory loss in patients. A transgenic mouse model of inducible olfactory inflammation under controlled expression of the microglia-derived proinflammatory cytokine TNF-α successfully showed that TNF-α can produce infiltration of inflammatory cells into the OE [62]. It is also reported that neural synaptic activity, as determined by electro-olfactogram (EOG) recordings, was altered in a TNF-α concentration-dependent manner; indeed, the EOG response was almost abrogated upon long-term induction of TNF-α, while it recovered to a normal level after TNF-α withdrawal. Moreover, TNF-α can suppress regenerative capacity in the OE by inhibiting the proliferation of neural progenitors and impeding the basal turn-over rate of mature olfactory neurons [62].
The contribution of microglia-derived proinflammatory cytokines in the olfactory system has been also described in bacterial infection models [5294]. In these studies, mice were challenged with Staphylococcus aureus after Triton X-100 (Sigma-Aldrich, USA) treatment in the ipsilateral nasalis. Infection of S. aureus led to a significant olfactory damage accompanied by rapid increases in proinflammatory cytokines IL-6 and TNF-α. In addition, inducible nitric oxide synthase-expressing microglia were accumulated in the OE and OB in response to bacterial exposure [52]. Although the physiological meaning of microgliosis following bacterial infection has not been fully described, microglia can play key roles in the development and resolution of inflammatory responses in the olfactory system [87].
Neurodegenerative models (Niemann-Pick disease type C and other diseases)
Using simple but reliable behavior tests reveals that rodents bearing neurodegenerative conditions usually show the distinctive signs of olfactory loss similar to those of humans (Table 1). AD-affected transgenic mice are the most widely studied animal model for olfactory loss, and most related papers have focused on olfactory memory loss or learning ability defects because AD is mainly associated with defective hippocampal cognitive CNS functions in humans [95108110]. In the OB, neuronal death and neurite abnormality have been observed with cholinergic- and noradrenergic signaling defects, leading to olfactory dysfunction in AD mice [51]. It has been also noted that amyloid beta deposits were found in the AD-affected OB early in life and accumulated throughout the olfactory system with age [100]. Defective neuronal diversity with decreased neurogenic activity in the OB has been also reported in other neurodegenerative disease mice as summarized in Table 1. These findings have indicated the contribution of neuronal abnormality to the development of olfactory loss in neurodegenerative diseases, although little has been reported about the molecular mechanism(s) underlying neural defects in the olfactory system.
In general, microgliosis is a common pathological feature of neurodegeneration throughout the brain, and abnormal microglial activation in the OB is clearly observed in AD- and PD-affected patients [37]. However, contribution of microglia to olfactory defects has been somewhat underestimated because microgliosis is often regarded as an inevitable outcome of autonomous neural damage. In this regard, we have studied the correlation between olfaction and microgliosis by using mice models for Niemann-Pick disease type C (NPC), a rare but fatal neurodegenerative disorder, to elucidate the neuropathological effects of microglial activity on olfactory loss during the neurodegenerative process.
NPC is an autosomal recessive, incurable disease found in approximately one among 120,000 live births [96]. Ninety-five percent of NPC cases result from genetic mutations in the NPC1 gene, referred to as NPC type C1 (NPC1), while the remaining 5% of cases originate from the NPC2 gene mutation, referred to as NPC type C2 (NPC2) [74]. Both NPC1 and NPC2 proteins are associated with the process of cholesterol egress in late endosomes and lysosomes, although the exact mechanism of NPC1/2-mediated cholesterol trafficking has not been fully elucidated [90]. Because NPC1/2 proteins are ubiquitously expressed in mammalian cells, NPC is considered a neurovisceral disorder, and its pathologic signs are easily observed in various organs including liver, spleen, and lung, but the CNS is the most affected region [9096]. Importantly, the severity of neurological symptoms is regarded as the most decisive factor for the determining the prognosis of NPC because neurodegeneration is ultimately responsible for the death of most patients. Indeed, delayed development of neurological symptoms seems to be correlated with longer survival in patients [4453]. Therefore, therapeutic strategies for NPC management are mainly focused on the early detection, prevention, and amelioration of neuropathologic signs of patients [73].
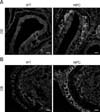
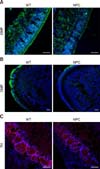
To date, noticeable sign of an olfactory defect in NPC patients have not been reported; however, growing evidence collected from animal studies has suggested the presence of olfactory dysfunction in NPC, as has been observed in other neurological disorders. Hovakimyan et al. [54] reported that electrophysiological activity of olfactory mucosa derived from the OE was significantly decreased in response to various odor stimuli in NPC mice compared to the activity level in the wild-type counterpart. More direct evidence of olfactory dysfunction in NPC mice was obtained from the results of buried food finding tests. During those tests, normal mice successfully located the hidden food within the prescribed time limit (3 min) while NPC mice had difficulty in carrying out the task, showing a high failure rate, thus implying that olfaction is impaired in NPC status mice [83]. Histologically, filipin staining revealed massive accumulations of cholesterol in the principal components of the olfactory system, the OE and OB (Fig. 1). Furthermore, the numbers of mature OSNs and periglomerular neurons were profoundly decreased in these regions, partially due to the reduced survival of precursors for OSNs and OB neuroblasts (Fig. 2) [83], supporting the suggestion that axodendritic synaptic activity between OE and OB might be insufficient for normal processing of olfactory information in NPC mice. Interestingly, abnormal microglial activation was observed in the NPC-affected OB, and the anti-inflammatory agent cyclosporin A (CsA) successfully prevented abnormal microgliosis and immature neuronal apoptosis in the NPC-affected OB, leading to the recovery of olfaction [83].
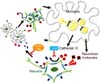
Activated microglia can perform multiple functions including cytokine production, phagocytosis, and antigen presentation by interacting with neighboring neurons via the production of several soluble factors to create coordinated neuroinflammatory or neuroprotective responses depending on the context [2949]. Therefore, we extended our previous studies by focusing on microglia-neuronal crosstalk to elucidate the underlying mechanisms of the massive microgliosis in the NPC-OB. It was noted that the olfactory deficit in NPC mice seems to be, in part, the result of bi-directional microglia-neuron interaction [82]. In detail, a higher level of neuronal chemokine Cx3cl1, which mediates neuronal-microglial communication through binding to its specific receptor CX3CR1 on microglia, was observed in NPC mice-derived cerebrospinal fluid (NPC-CSF) than in wild-type mice. Moreover, most Iba1-positive amoeboid NPC1 microglia expressed the Cx3cr1 marker, implying a correlation between microglial activation and Cx3cl1-Cx3cr1 signaling. To determine the biological and pathological significance of our results, we neutralized Cx3cl1 function with a specific Cx3cl1 antibody and then undertook histological analysis and behavioral tests. Importantly, blockage of Cx3cl1-Cx3cr1 interaction prevented neurotoxic microglial behavior and was followed by increased survival of OB neurons and partial recovery of olfactory loss in NPC1 mice. Moreover, upregulation of Cx3cl1-Cx3cr1 signaling was partially mediated by the catalytic activity of microglial cathepsin S (Ctss) because the nasal infusion of the Ctss inhibitor LHVS could successfully suppress microglial activation by reducing Cx3cl1 secretion into NPC-CSF. As a result, LHVS not only prevented OB neuronal damage, but it also improved the olfactory function of NPC1 mice. Finally, we suggested a causal link between NPC1 dysfunction and abnormal Ctss activity of NPC1 microglia; in detail, disturbed cholesterol homeostasis caused by NPC1 mutation led to cholesterol accumulation in the lysosome and, in turn, activated p38 MAPK signaling, resulting in Ctss activation and maturation (Fig. 3) [82]. Interestingly, the transcription level of Ctss was reported to be increased in the liver and cerebellum of NPC-affected patients [2], implying that the results from the mouse experiment could be applicable in clinical fields.
Summary
In this review, we have pointed out the possible contribution of microglia to olfactory impairment and, based on our recent studies with NPC mice, emphasized the usefulness of animal models in this field. In NPC status, the integrity of the central olfactory system is disrupted due to a cholesterol accumulation, causing excessive crosstalk between neurons and microglia via the Cx3cl1-Cx3cr1 axis and leading to olfactory dysfunction. Moreover, the application of specific inhibitors for the target molecule indicates therapeutic potential for olfactory recovery in NPC mice. Therefore, animal study can provide not only a thorough insight into understanding the nature of olfactory system, but also, it can inspire the development of novel therapeutic strategies for olfactory loss in humans.

 XML Download
XML Download