

 PDF
PDF Citation
Citation Print
Print


The life histories of vector-borne and/or trophically transmitted pathogens involve complex associations with a wide range of environmental factors including abiotic/biotic components and the ecology of both vector and host species, thus making pathogens particularly vulnerable to environmental conditions [14]. Therefore, attempts to elucidate variation in interactions between animal hosts and their pathogens with complex life cycles should consider the potential effect of different habitat conditions [14]. In this study, we investigated the potential association between habitat type (urban and rural) and prevalence of feral domestic cats (Felis catus) infected with three pathogens that have different transmission modes: Toxoplasma (T.) gondii gondii, Bartonella (B.) henselae, and hemoplasma. T. gondii is a zoonotic protozoon for which only felids, such as domestic cats, serve as definitive hosts [7]. Globally, domestic cats are distributed throughout a wide range of habitat types, and their interaction with pathogens is expected to vary in response to their surroundings, from sites in highly urbanized cities to those in protected natural areas [4]. Domestic cats can be infected with T. gondii trophically through predation of an infected intermediate host, such as a rodent, or through environmental transmission-contact with oocysts in fecal matter or contaminated abiotic materials such as soil and vegetation [7]. B. henselae is a flea-borne bacterial pathogen, and feline species, such as domestic cats, can serve as a reservoir host [1]. Mycoplasma haemofelis (Mhf) and Mycoplasma haemominutum (Mhm) are important bacterial pathogens (hemoplasmas) in feline species and may cause mild to severe anemia, especially when cats are co-infected with feline immunodeficiency virus and/or feline leukemia virus [15]. It is assumed that hemoplasmas are transmitted through either direct contact with saliva (e.g., biting) or blood of infected individuals (e.g., arthropod-borne) [15]. We hypothesized that the prevalence of two vector-borne pathogens (B. henselae and hemoplasma) would be higher in rural habitats due to frequent exposure of feral cats to potential vector species, such as ticks and fleas. We also expected a higher prevalence of a primarily trophically transmitted parasite (T. gondii) in feral cats from rural populations due to higher predation rates of rural feral cats on the parasite's intermediate host.
We collected samples from feral cats (n = 117) in five different districts within a metropolitan area (Seoul, Korea) and in four rural areas in Korea. Seoul (37°33′59.53″N 126°58′ 40.69″E) is an intensively developed city, with a human population density of 17,000 per square kilometer [9]. All five urban trapping sites were selected among residential areas densely packed with multi-story residential buildings, one of the most typical housing structures in Seoul according to data from the Seoul Metropolitan Government. Rural sites were selected among areas with typical agricultural landscapes composed of extensive rice paddies, shrubland, and sparsely dispersed houses. The rural sites sampled were in four different regions, Yangpyeong county (37°46′15.262″N 127°42′10.875″ E), Cheorwon county (38°11′16.6″N 127°16′48.8″E), and the outskirts of Gwangju city (37°25′16.3″N 127°19′14.1″E) and Chuncheon city (37°59′37.0″N 127°43′50.7″E).
One hundred seven feral cats were live-trapped in the summers of 2013 and 2015 by using ten Tomahawk traps (Tomahawk; Live Trap, USA). Ten additional samples from cats in a district within the Seoul city were collected from a ‘Trap-Neuter-Return’ center, thus providing a total of 117 samples for analysis. Male cats comprised approximately 59.8% (70/117) and female cats approximately 40.2% (47/117) of the sampled cats. The ratio of male to female cats was not significantly different between the two habitat types (Chi-squared test; p = 0.39). DNA was extracted from whole-blood samples by using the QIAamp DNA mini kit (Qiagen, Germany), and was used to detect hemoplasma infection through a subsequent PCR assay. We amplified a partial segment of the 16S rRNA gene by using a PCR protocol described in a previous study [3]. Amplified products were separated by electrophoresis on a 1.2% agarose gel. PCR amplicons from the positive samples were sequenced to identify the nature of the amplified product. The sequence homology of the sequenced 16S rRNA gene from the positive samples was examined by using the BLAST program (National Center for Biotechnology Information; USA). Serological assays for T. gondii and B. henselae were performed by using commercial diagnostic kits and following the protocols provided by the manufacturers. The indirect immunofluorescence assays (IFA) were performed by using the Bartonella henselae Feline IFA IgG Kit (Fuller Laboratories, USA) for B. henselae assays; the IFA antigen slides were produced by using B. henselae co-cultivated with Vero cells. Serum dilutions from 1/64 were applied to antigen slides. A 1/50 dilution of fluorescein-conjugated goat anti-cat immunoglobulin G (IgG; heavy and light chains) in 0.01% Evans blue was used as the conjugate. A known positive sample was provided by the manufacturer. Only specimens with unambiguous bright fluorescence in a dilution of 1/64 or greater were considered seropositive for B. henselae. For the T. gondii assays, we undertook point-in-care ELISA with a SensPERT Feline Toxo IgG Ab Test Kit (VetAll Laboratories, Korea) with sensitivity of 98% and specificity 98.9% compared to IFA for IgG.
Statistical analyses for the association between the prevalence of tested pathogens, habitat type, and sex were performed by applying Chi-squared tests. Phi-coefficient (φ) was calculated to show the effect size of the association between variables. The relative risk and the 95% confidence interval for pathogen infection were calculated for each pathogen. All statistical analyses were performed by using the statistical software R. Ethical approval for the use of animals in this study was obtained from the Institutional Animal Care and Use Committees of the University of Georgia, USA (IACUC No. A2013 04-009-Y1-A0) and Seoul National University, Korea (SNUIBC-R131118-1). To compare the relationship among the hemoplasma sequences identified in this study with other sequences deposite in GenBank, sequences were aligned by using the Clustal W algorithm [18] and analyzed by using the program MEGA 6 [16]. Phylogenetic trees were constructed by using the neighbor-joining method and the dataset was resampled 1,000 times to generate bootstrap values.
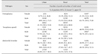
Among the 117 cats, 56 cats (47.9%) were PCR-positive for hemoplasma infection. Serological detection of T. gondii and B. henselae antibodies (IgG) was performed in 112 of the 117 cats, and 56 cats (50%) and 40 cats (35.7%) were positive for T. gondii and B. henselae antibodies, respectively. Additional hemoplasma, T. gondii, and B. henselae occurrence results by habitat type and sex are summarized in Table 1. Significant associations between pathogen prevalence and external factors (i.e., habitat type and sex) were only observed between B. henselae and habitat type, and between hemoplasma and sex. Occurrence of B. henselae was higher in urban areas (Chi-squared test; p = 0.03), while occurrence of hemoplasma was higher in male cats than in females (Chi-squared test; p = 0.00). Additional information regarding the associations are summarized in Table 2. Among the hemoplasma-positive samples, we detected ‘Candidatus Mycoplasma turicensis (CMt)’ sequences in two cat samples. The identified CMt sequences were submitted to GenBank (accession Nos. KY046311 and KY046312). The phylogenetic tree of the CMt sequences and additional sequences of Mhf and Mhm identified in this study (GenBank accession Nos. KY432678–KY432680) are depicted in Fig. 1.
For parasites relying on trophic transmission, such as raccoon roundworms, lower host infection prevalences have been reported in urban habitats [813] than in rural conspecifics, presumably due to a lower predation rate on prey species serving as intermediate hosts in urban areas. However, unlike parasites that solely rely on consumption of intermediate hosts by definitive hosts for transmission, T. gondii can also be transmitted through direct contact via environmental contamination with T. gondii oocysts. It is possible that the relatively higher seroprevalence in urban cats in this study was due to a high population density of urban feral cats and more frequent exposure of cats to T. gondii oocysts through environment contamination, which compensated for the reduced trophic transmission of T. gondii. Considering the behavior of feral cats, such as defecating in soil and burying fecal matter, the apparently higher T. gondii transmission in urban cats presents potential risks in urban playgrounds and/or parks of T. gondii dissemination, and warrants further investigation of T. gondii transmission ecology in urban habitats [5].
Seroprevalence of B. henselae was significantly higher in urban feral cats. Previously, prevalence of vector-borne pathogens or infestation of vector species has been reported to be higher in rural-dwelling host populations than in urban populations [212]. However, variation in such a pattern may exist, depending on the ecology of the vector species involved in the transmission. For instance, the buffered microclimate of urban buildings can provide an ideal condition for flea populations. Although the dry and high temperatures of urban areas may have a negative effect on the survival of fleas [10], indoor habitats, such as abandoned buildings, may provide environments where flea populations can grow throughout the year [11]. Considering the lack of data on flea infestation rates or prevalence of other ectoparasites in urban feral cats in Korea, additional studies evaluating flea infestation rates and flea-borne pathogens in urban feral cats are required to clarify the effects of habitat type on flea epidemiology.
We did not observe significant habitat-related differences in hemoplasma prevalence. However, a significant sex difference was observed, with males showing higher prevalence in both habitat types (urban males: 60%; rural males: 74.2%). This result is similar to those in previous studies and is potentially explained by bite-related transmission between male cats due to intraspecific aggression [6]. Furthermore, the prevalence of hemoplasma in females between the two habitat types was also significantly different, with females from rural areas showing approximately four-times greater prevalence than that in urban areas (urban females: 8.7%; rural females: 32%). Considering that feline hemoplasma may also be transmitted through blood-sucking arthropod vectors in addition to salivary transfer [1517], the pattern of hemoplasma prevalence between urban and rural female cats may be the combined result of two transmission modes (vector-borne and saliva through biting). To best of our knowledge, this is the first report of CMt in domestic cats in Korea, which indicates the need for further studies into its pathogenicity and its interactions with host and other hemoplasma species.
Feral cats are hosts of a number of zoonotic pathogens, including T. gondii and B. henselae, and many of them have complex life histories and multiple transmission modes. Based on the results of this study, associations between habitat type and host-parasite interactions may vary in relation to the transmission mode(s) of each pathogen. With rapid urbanization, interactions between humans and urban feral cats, and potentially other urban wildlife, are raising diverse social issues, including public health issues. Hence, further investigations into the ecological and epidemiological traits of urban feral cats, such as infestation rates of ectoparasites and dietary content of feral cats in different habitat types, will provide critical data required to undertake appropriate preventative measures related to public health and epidemiological management of feral cat populations.


 XML Download
XML Download