

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
In April 2009, emergence of the novel H1N1 2009 (pH1N1) virus originating from swine initiated the first human pandemic of the 21st century [23]. This virus continued to spread among human populations and was transmitted to other animal species such as domestic pigs [18,25], turkeys [12], and cats [1]. Recent evidence has shown that novel pH1N1 viruses were generated by reassortment events between pH1N1 and endemic swine H1N1 (swH1N1) viruses in pigs in many countries, including Thailand [9]. It is known that influenza A virus (IAV) transmission between humans, pigs, and avian species is common [29]. Thus, these novel pH1N1 reassortants generated in pigs that already contained avian-like genes have a high potential for transmission back to humans and birds. In addition to pigs, quails [Coturnix (C.) japonica or C. coturnix] are another known species capable of being mixing vessels for IAV as they express both α-2,3- and α-2,6-linked receptors [30]. Quails are broadly susceptible to infection with several subtypes of both mammalian and avian influenza viruses [11]. However, little information is available on the susceptibility or pathogenicity of pH1N1 and swH1N1 viruses in quails.
The quail is a land-based bird commonly raised by humans worldwide. The first IAV outbreak in quails was reported in Italy in the late 1960 [15]. Subsequent studies showed that quails could be naturally infected with numerous influenza subtypes of avian, human, and swine origins, such as the H3, H4, H5, H6, H7, H9, and H10 subtypes of avian influenza viruses as well as human H1N1 and swine H3N2 influenza viruses [10,16]. In addition to quails naturally infected with IAVs, ones experimentally infected with the highly pathogenic avian influenza (HPAI) H5N9 virus show no clinical signs while shedding the virus [28]. However, the virus can be transmitted to chickens, leading to death [28]. Some studies suggested that quails are more susceptible than chickens to experimental infection with the HPAI H5N1 viruses isolated from southeastern China and Thailand [22,31]. It was demonstrated that quails experimentally infected with H5N1 viruses isolated from chickens have longer survival period than chickens [3,22]. Previous experimental infection studies also showed that quails are broadly susceptible to 14 subtypes of avian influenza viruses as well as swine influenza viruses (classical H1N1, H3N2, and H1N2) and human-like H1N1 virus [11,19].
Recent studies have described the pathogenicity of pH1N1 virus in many mammalian species including mice, ferrets, non-human primates, and pigs [6]. Other studies showed that the virus does not result in disease in most tested poultry species, but it can replicate and be shed with limited transmission among quails [5,27]. However, few reports are available on the pathogenicity of pH1N1 in quails, and direct comparison of pH1N1 pathogenicity with other swine and avian influenza viruses has not been made. In the present study, the pathogenicity, shedding, and transmission characteristics of pH1N1 virus in quails were investigated. These characteristics were compared to those of endemic Thai swH1N1 virus and the low pathogenic avian influenza virus (LPAIV) Thai duck H3N2 (dkH3N2) virus.
Materials and Methods
Viruses
Three viruses of different IAV subtypes, pH1N1 (A/swine/Thailand/CU-RA4/2009), swH1N1 (A/swine/Thailand/CU-CB1/06), and LPAIV duck H3N2 (A/duck/Thailand/AY-354/08) isolated from pigs and ducks in Thailand, respectively, were used in this study. The swH1N1 and dkH3N2 viruses belonged to the classical Eurasian H1N1 lineage or Eurasian avian lineage, respectively. pH1N1 and swH1N1 viruses were propagated by four passages in Madin-Darby canine kidney (MDCK) cells (ATCC, USA) as previously described [8]. The dkH3N2 virus was propagated once in the allantoic cavity of 9-day-old embryonated chicken eggs [25]. The viruses were harvested from cell culture superanatant of pH1N1 and swH1N1 infected MDCK cells and from allantoic fluid of dkH3N2 inoculated embryonated chicken eggs. The viruses were clarified by centrifugation at 5,000 g for 15 min, 4℃. Next, 50% tissue culture infectious doses (TCID50) of the viruses were made as previously described by Reed and Muench [21] following serial titration in MDCK cells [24]. All virus stocks were diluted in minimal essential medium (MEM) (Invitrogen, USA) to 106 TCID50/mL. Virus propagation and handling were performed in biosafety level (BSL) 2 containment facilities.
Animals
Animal experiments were performed in the BSL-3 containment facility of the Faculty of Veterinary Science, Chulalongkorn University (Thailand). Animal protocols were conducted in accordance with the guidelines and approval (approval No. 0931048) of the Institutional Animal Care and Use Committee at Faculty of Veterinary Science, Chulalongkorn University (Thailand). Seventy-five 4-week-old quails (C. coturnix) were used in the study. The source of quails was from a local farm in Thailand that has been proved to be free from many pathogens, including influenza A viruses. Prior to viral infection, all quails were confirmed to be serological negative with a hemagglutination inhibition (HI) assay specific for pH1N1, swH1N1, and dkH3N2 viruses as described below, and an anti-influenza A NP-ELISA (Idexx Laboratories, USA) according to the manufacturing protocol. In addition, oropharyngeal (OP) and cloacal (CL) swabs were collected and tested for IAV RNA prior to inoculation to ensure an absence of IAV infection. Oropharyngeal (OP) and cloacal (CL) swabs were tested for IAV RNA by performing RT-PCR assay using primers that specific for M gene. All IAV-infected quails were housed in biosafety cabinet (class III) isolators (Ingénia Technologies, Canada) ventilated under negative pressure with HEPA-filtered air. The feed specific for quails (Betagro, Thailand) and water was provided ad libitum.
Experimental design
Three groups of 15 birds each were inoculated by dripping 106 TCID50/mL of pH1N1, swH1N1, or dkH3N2 virus in a total volume of 0.25 mL orally (0.2 mL) and intranasally (0.05 mL). An additional 15 naive quails were mock-inoculated with 0.25 mL of MEM and served as the negative control group. To monitor virus transmission, five IAV-seronegative quails were introduced to each group 1 day post-inoculation (DPI) to allow direct contact. Clinical signs, including depression, ruffled feathers, diarrhea, and respiratory distress, were monitored daily for 7 DPI. OP and CL swabs (Copan Diagnostics, USA) were collected daily for 7 DPI. Swabs were suspended in viral transport medium [MEM containing 2% bovine serum albumin (Invitrogen, USA) and 100 µg/mL of gentamicin (Invitrogen, USA)] and stored at -80℃ until virus shedding was evaluated. On 3, 5, and 7 DPI, five quails from each group were humanly euthanized by pentobarbital injection via intravenous route for gross lesion examination. Tissue samples, including ones from the brain, heart, trachea, lung, spleen, liver, intestine, pancreas, and kidney, were collected for histopathology and immunohistochemistry (IHC) examinations. All of the remaining inoculated and contact quails from each group were bled at 7 DPI by 22 gauge needle with 3 mL syringe. One mL of blood was centrifuged at 2,500 g for 10 min for further serological analysis.
Virus isolation
In order to evaluate the virus shedding patterns, virus isolation was performed until 7 DPI from OP and CL swabs for the quails that remained. Virus was titrated in MDCK cells as previously described [8]. Briefly, 100 µL of swab solutions serially diluted 10-fold in MEM (Invitrogen, USA) containing 0.3% bovine albumin fraction V (Invitrogen, USA) and 1 µg/mL tosylsulfonyl phenylalanyl chloromethyl ketone-treated trypsin (Sigma-Aldrich, USA) were used to inoculate MDCK cells in 96-well culture plates (Thermo Scientific, USA) and incubated at 37℃ in 5% CO2 for 48 h. Virus identification was performed by staining with an anti-IAV nucleoprotein monoclonal antibody (clone HB-65; ATCC, USA) followed by horseradish peroxidase-conjugated rabbit anti-mouse IgG (Dako, Denmark) [24]. Color was developed using a chromogen 3-amino-9-ethylcarbazole substrate (Sigma-Aldrich, USA) [24]. Each test included mock-infected negative control cells and positive control cells infected with a known titer (106 TCID50/mL) of virus. The virus titers in each swab were expressed as log10TCID50/mL and determined using the method by Reed and Muench [21].
Histopathology
Tissue samples collected from quails 3, 5, and 7 DPI were fixed at room temperature in 10% neutral buffered formalin solution for a minimum of 48 h. After fixation, the tissues were immediately embedded in paraffin and cut by microtome (Sakura Finetek, Japan) into 5-µm sections. For the histological examination, the sections were stained with hematoxylin and eosin and assigned a score of 0 to 3 based on the degree of inflammation by a veterinary pathologist (R.T.): 0, no lesion; 1, mild inflammation characterized by focal infiltration of inflammatory cells; 2, moderate inflammation characterized by multifocal infiltration of inflammatory cells; 3, severe inflammation characterized by diffuse infiltration of inflammatory cells.
IHC
The presence of IAV-specific antigen in the tissues collected was identified by IHC as previously described [24]. In brief, duplicate 5-µm sections were incubated at 4℃ overnight with a mouse anti-IAV monoclonal antibody clone EVS 238 at dilution 1 : 300 (HB65 like; BV European Veterinary Laboratory, the Netherlands) followed by incubation for 45 min at room temperature with EnVision polymer reagent (Dako, Denmark). Immunohistochemical signals were visualized using 0.4 mg/mL 3,3-diaminobenzidine tetrahydrochloride (Sigma-Aldrich, USA) by incubating for 2 min at room temperature. Sections were then counterstained in hematoxylin. Each test included a positive and negative control.
Serological analysis
Serum samples collected from each quail at 0 and 7 DPI were treated with 50 mg/mL receptor destroying enzyme (Seiken, Japan) for 18~20 h at 37℃ and tested with an HI assay for the presence of specific antibodies against the homologous virus used for inoculation as previously described [8]. The HI assay was performed using 0.5% v/v chicken red blood cells (CRBCs) in phosphate buffered saline. The HI titer was expressed as the reciprocal of the last dilution that contained CRBCs with no agglutination, and reported as the geometric mean. Negative and positive controls were included in the assay. None of the quails had HI titers against pH1N1, swH1N1, or dkH3N2 viruses prior to inoculation. Thus, samples collected after exposure to the viruses with a titer ≥ 10 were considered to be seropositive [2].
Statistical analysis
Significant differences in virus shedding titers and histopathological lesion scores were evaluated with an analysis of variance (ANOVA) or a non-parametric Wilcoxon/Kruskal-Wallis test (rank sum test), respectively, using JMP 5.1 software (SAS, USA). p-values < 0.05 were considered to be statistically significant.
Results
Clinical observations
No severe clinical signs or mortality were observed in any of the IAV-inoculated (n = 45), contact (n = 15), or negative control (n = 15) quails during the 7 days of observation. However, six of 15 quails inoculated with pH1N1 and four of 15 quails inoculated with dkH3N2 had mild nasal discharge observed during necropsy.
OP and CL shedding
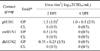
All of the three tested viruses were detected predominantly in the oropharynx (from OP swabs) of the inoculated quails (pH1N1 = 4/5, swH1N1 = 1/5, and dkH3N2 = 3/5) but were detected in the cloaca (from CL swabs) in only a small number of the inoculated quails (pH1N1 = 2/5, swH1N1 = 2/5, and dkH3N2 = 2/5) with lower mean virus titers compared to the oropharynx (Table 1). Four out of five pH1N1-inoculated quails shed the virus oropharyngeally from 1 DPI and up to 5 DPI; one bird shed virus until 7 DPI (Table 1). Quails inoculated with pH1N1 virus showed similar virus shedding trends compared to dkH3N2-inoculated birds, which shed slightly higher mean titers of the virus on the OP swabs for a shorter period (1.3 to 3 log10TCID50/mL from 1 to 5 DPI) compared to quails inoculated with pH1N1 (1.3 to 2 log10TCID50/mL from 1 to 7 DPI). However, this difference was not statistically significant. Three out of five dkH3N2-inoculated quails shed virus oropharyngeally 1 DPI; one bird shed virus up to 5 DPI while the remaining quail shed virus for only 2 or 4 days (Table 1).
In contrast to the marked shedding observed in pH1N1- and dkH3N2-inoculated quails, only one out of five quails exposed to swH1N1 shed virus at very low titers (0.5 log10TCID50/mL) for 4 days through the OP route (Table 1). CL shedding from all groups was observed in a smaller number of quails (20 to 40%) compared to OP shedding (20 to 80%) for 2 or 3 days with low titers of virus (0.5 to 1.5 log10TCID50/mL) except one swH1N1-inoculated quail that showed a CL shedding titer of 3 log10TCID50/mL 5 DPI (Table 1). No virus was isolated from swabs of the negative control quails. Overall, quails inoculated with pH1N1 or dkH3N2 shed higher amounts of virus in the OP swabs compared to birds inoculated with swH1N1.
Transmission
Two out of five pH1N1 contact quails shed virus at low virus titers (1 to 1.5 log10TCID50/mL) from the oropharynx 2 and 4 DPI. Low levels of swH1N1 (0.5 log10TCID50/mL) or dkH3N2 (0.75 log10TCID50/mL) virus were isolated in OP swabs 2 DPI in one out of five and two out of five contact quails, respectively. Higher levels of virus shedding for a longer period of time were detected in the OP swabs of pH1N1 contact quails (1 to 1.5 log10TCID50/mL at 2 and 4 DPI) compared to swH1N1 and dkH3N2 contact quails (0.5 to 0.75 log10TCID50/mL at 2 DPI). Virus was not detected in CL swabs collected from any contact quails in all groups (Table 2). Overall, this result revealed that all of the three tested viruses were poorly transmitted among the quails.
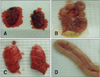
Gross pathology
No gross lesions were observed in tissues from swH1N1-inoculated quails, contact quails in any group, or negative control quails. Gross lesions were detected mainly in the lungs and intestinal tissues of quails inoculated with pH1N1 and dkH3N2. Lesions included congestion of the lung and duodenum (Fig. 1). Extent and degree of severity of the gross lesions in the lung and intestinal tissues appeared to be the same at 3, 5, and 7 DPI when comparing the pH1N1- and dkH3N2-inoculated groups. The number of pH1N1-inoculated quails with gross lung lesion gradually increased over 3, 5, and 7 DPI (3 DPI = 2/5, 5 DPI = 3/5, and 7 DPI = 5/5). In contrast, all dkH3N2-inoculated quails (5/5) exhibited lung lesions at all time points. Intestinal lesions were observed in some pH1N1-inoculated quails at all time points (3 DPI = 2/5, 5 DPI = 2/5, and 7 DPI = 4/5) whereas these lesions were observed in all dkH3N2-inoculated quails (5/5) only on 7 DPI (Table 1).
Histopathology and IHC
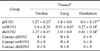
Histopathological lesions in tissues from quails inoculated with pH1N1, swH1N1, and dkH3N2 were mostly restricted to the trachea, lung, and duodenum. No significant lesions were observed in the remaining tissues of any inoculated quail. Heterophilic-to-lymphocytic tracheitis, deciliation and sloughing of epithelial cells in the trachea, mild to moderate bronchitis, peribronchiolar cuffing characterized by heterophilic and lymphocytic infiltration surrounding the bronchioles, mild to moderate diffuse pulmonary congestion and hemorrhage, and mild to moderate duodenitis with inflammatory cells infiltration (such as heterophils and macrophages) were the most commonly observed lesions (Fig. 2). These were similar among all inoculated groups at all time points, but the most severe lesions were detected 7 DPI. Quails inoculated with pH1N1 and dkH3N2 had significantly higher mean histopathological scores for the duodenum than birds in the swH1N1 group (p < 0.05; Table 3). No histopathological lesions were observed in any of the tested organs from contact quails in all groups or the negative control quails. Collectively, these findings demonstrated that pH1N1 and dkH3N2 infection caused more histopathological lesions in quails than swH1N1.
Minimal staining for IAV nucleoprotein antigen was observed in macrophages within the duodenal lamina propria of one quail inoculated with dkH3N2 at 3 DPI (Fig. 2). No IAV antigen was detected by IHC in any tested organs from all groups of quails.
Serology
Serum collected from each quail 0 and 7 DPI were analyzed for the presence of homologous virus-specific antibodies using an HI assay. All quails were seronegative for pH1N1, swH1N1, and dkH3N2 virus prior to inoculation. Quails in the negative control group remained seronegative for all viruses throughout the study. Two out of five pH1N1-inoculated quails had HI antibody titers of 10 and 320 against homologous virus 7 DPI (Table 4). HI antibodies were not detected in any pH1N1 contact quails. Three out of five quails inoculated with swH1N1 had low levels of HI titers (10) 7 DPI. Four out of five swH1N1 contact quails produced low levels of HI titers (10) 7 DPI (Table 4). In contrast, seroconversion was not detected in any of the dkH3N2-inoculated and corresponding contact quails.
Discussion
Previous observations indicated that quails are a potential intermediate host for generating novel reassortant IAVs with pandemic potential [30]. With the emergence of the pH1N1 virus, concerns about novel highly pathogenic reassortant pH1N1 generated in quail necessitated detailed studies of pH1N1 infection in this species. A few reports are available on susceptibility of quails to pH1N1 infection [5,27]. However, those investigations were restricted to reporting only virus shedding and did not offer direct comparison with IAVs endemically circulating in other species. In the present study, the susceptibility and pathogenicity of pH1N1 in quails were determined and directly compared to that associated with swine (swH1N1) and avian (dkH3N2) influenza viruses. We confirmed that the quails were susceptible to pH1N1 infection that led to a prolonged virus-shedding period without the development of any clinical signs. Viral pathogenesis was restricted to the respiratory and intestinal tracts but the main mode of virus transmission to contact birds was through an OP route. Importantly, our study revealed that the quails were more susceptible to infection with pH1N1 and dkH3N2 compared to the endemic swH1N1 strain.
As observed in previous studies [5,11,19,27], the pH1N1 swine influenza virus and LPAIV did not produce clinical signs in either the infected and contact quails. Furthermore, our results were in agreement with ones from previous studies demonstrating that quails inoculated with pH1N1 are asymptomatic but shed virus mainly from the respiratory tract [5,27]. This finding corresponded to data from another study showing high expression of α-2,6-linked receptors in quail trachea [30]. In addition, our data agreed with those of previous investigations showing that LPAIV- and swine influenza virus-infected quails do not develop clinical signs, and shed virus predominantly from the respiratory tract that is poorly transmitted to contact quails [10,11,19]. In the present study, low or undetectable levels of virus shedding from the cloaca were observed in quails infected with all three viruses, indicating that these viruses replicated more efficiently in the respiratory tract than in the intestinal tract.
Our results showed that quails inoculated with pH1N1 could shed virus from the oropharynx for up to 7 DPI. This finding was different from those of previous studies indicating that quails can shed pH1N1 for only 4 or 5 days [5,27]. However, the levels of virus shedding were slightly higher compared to those observed in our study. This difference might be related to the strain of pH1N1, route of inoculation, and viral doses. The swH1N1 used in our study was classified as an avian-like virus since most of its genes belonged to an avian-like swine lineage. However, lower levels of OP shedding were detected in quails inoculated with swH1N1 compared to birds exposed to classical swH1N1 virus as previously reported [11]. Moreover, our study demonstrated that lower levels of dkH3N2 from a Eurasian lineage were shed from the oropharynx of infected quails compared to Eurasian H3 virus previously described in the literature [10,11]. Another study showed that mammalian H1 viruses, including pH1N1 and swH1N1 viruses, have a typical human-like preference for the α-2,6-linked receptor [5]. However, swH1N1 replicates poorly in quail respiratory tract whereas pH1N1 could replicate more efficiently despite the presence of α-2,6-linked receptors in quail trachea. Therefore, the limited replication of swH1N1 compared to pH1N1 could not be explained based only on receptor distribution. This might be due instead to a combination of internal pH1N1 genes not shared with swH1N1 that may have enabled pH1N1 to replicate in the quail respiratory tract. Overall, our study showed that quails inoculated with pH1N1 and dkH3N2 shed higher virus titers in OP swabs compared to birds inoculated with swH1N1. However, levels of virus were relatively low compared to ones reported in previous studies [5,11,27]. These data confirmed that all three viruses examined in our study replicated at low levels in quails.
pH1N1, swH1N1, and dkH3N2 caused pathological changes mainly in the respiratory organs and intestines of infected quails. Both sites are known to be the primary site for LPAIV replication in avian species [13]. However, minimal IAV antigen staining was only observed in the intestinal tract of a quail inoculated with dkH3N2. This finding is in agreement with observations in many avian species in which a minimal or lack of IAV antigen staining in tissues from LPAIV-infected birds was observed [7,13,14,20]. In addition, histopathological lesions observed in this study were similar to those associated with LPAIV infection in many avian species [13,14,17,20]. Therefore, it can be assumed that these lesions may be related to IAV infection. Pathological changes in pH1N1-infected bird observed in our investigation were different from a previous study in which only heterophillic-to-lymphocytic rhinitis and positive IAV antigen staining within the mucosa of the nasal cavity were noted [27]. This difference might be attributed to the route of inoculation and the swine-adapted strain of pH1N1 used in this study. Overall, the results from our study showed that pH1N1 and dkH3N2 were more pathogenic in quails than swH1N1. However, our findings also indicated that all three viruses are of low pathogenicity in quails.
Serological results confirmed the seroconversion of quails infected with pH1N1 and swH1N1 viruses and the corresponding contact quails. Our study detected HI antibodies against pH1N1 in pH1N1-inoculated quails a week earlier than a previous study [27]. On the other hand, none of the dkH3N2-inoculated or contact quails seroconverted 7 DPI, although virus shedding and pathological changes were observed in dkH3N2-inoculated quails. A previous study [4] determined that HI assays using CRBCs fail to detect serum HI antibodies in ring-necked pheasants and chukar partridges infected with avian influenza A viruses (AIVs) unlike those using horse red blood cells (HRBCs). This is because AIVs prefer to bind to α-2,3-linked receptors, which are predominantly found on HRBCs [26]. Thus, the absence of HI titers in the dkH3N2-infected quails may have been due to the use of CRBCs in the HI test. In addition, quails may need more time to develop HI antibodies against dkH3N2.
In conclusion, the present study demonstrated that quails were susceptible to infection with pH1N1, swH1N1, and dkH3N2 viruses. The replication and pathogenicity levels of pH1N1 and dkH3N2 in quails were much higher than that of swH1N1. However, none of the viruses replicated efficiently in quails and caused only mild lesions, indicating that these viruses act as LPAIV in quails. Overall, our findings revealed that the quails were susceptible to infection with all three different virus lineages yet appeared tolerant to the disease caused by these viruses. These data indicate important roles of quails as a susceptible host for mammalian and AIVs, and an intermediate host of IAVs. Importantly, the possibility of quails transmitting IAVs to other hosts as well as quail producers (in the case of farm-raised birds) can easily be overlooked since these birds are essentially asymptomatic while shedding IAVs. Awareness of IAV interspecies transmission and continued monitoring of IAVs in quails is thus crucial for IAV control and pandemic preparedness.



 XML Download
XML Download