

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Recently, there have been several serious outbreaks caused by the highly pathogenic avian influenza virus (HPAIV) of the H5N1 subtype among poultry and wild birds that could be transmitted to humans. The potential of this virus to cause a pandemic of serious respiratory diseases is high and thus, it poses a significant threat to global human health [7,11,37].
In Korea, there were three outbreaks caused by the H5N1 HPAIV in 2003/2004, 2006/2007 [19,44] and 2008. During the second outbreak, seven poultry farms were affected (two broiler breeding farms, two layer chicken farms, two duck breeding farms, and one quail breeding farm) and the outbreak lasted for 15 weeks from November 22, 2006 through to March 7, 2007. The naturally infected chickens and quails suffered high mortality and exhibited clinical signs including depression and a decrease in food consumption. In contrast, the ducks showed no mortality but a drop in egg production was observed. Our phylogenetic analysis of the 2006/2007 Korean isolates revealed that all the viruses of this outbreak were closely related and belonged to the A/bar-headed goose/Qinghai/5/2005-like lineage [19].
Outbreaks of Qinghai-like H5N1 viruses also occurred in poultry and wild birds in over 50 countries in Asia, Europe and Africa [8]. It was shown that these viruses may be transmitted between migratory waterfowl and thus, they present an unprecedented threat in terms of initiating a pandemic [6,22,39,42].
However, the main route of transmission appeared to be through contaminated equipment or via infected birds in live bird markets and farms [32]. Pathogenicity tests on Qinghai-like viruses revealed that most of the replication occurred in multiple organs and caused systemic infections in ducks, mice, and chickens. Moreover, some wild bird species became ill or died after being inoculated with the Qinghai-like virus, A/whooper swan/Mongolia/244/05 [2,3]. With regard to the genetic features of the Qinghai-like viruses, most bore a Lys 627 mutation in the PB2 gene [5,8,19,20,22,40] that has been associated with increased virulence in mice [12,38].
In this study, we assessed the characteristics of A/Chicken/Korea/IS/06, which was isolated from the index case of the 2006/2007 H5N1 HPAIV outbreak in Korea. We experimentally infected chickens, ducks and quails and examined their clinical and serological responses. We also determined the extent and duration of viral shedding as well as the viral titers in various tissues.
Materials and Methods
Virus
The A/chicken/Korea/IS/06 (H5N1) virus was isolated from the index case of the 2006/2007 HPAIV outbreak in Korea. It was propagated in specific pathogen-free (SPF) embryonated chicken eggs. The virus titers in oropharyngeal and cloacal swabs, and tissues were determined as a means to tissue culture infective doses (TCID50) into chicken embryonated fibroblast (CEF) cells.
Animals
Three avian species, specifically, SPF white Leghorn chickens, Cherry Valley ducks and Japanese quails were used for this study. Each species was housed separately in an isolator, ventilated under negative pressure with HEPA-filtered air, and maintained under continuous lighting. Appropriate food and water was provided ad libitum. All the animal experiments were conducted in bio-safety level 3 containment facilities, and all personnel were required to use respiratory protection when working with live viruses or the experimentally infected animals.
Experimental designs
Intravenous pathogenicity test: The intravenous pathogenicity tests with chickens and ducks were performed according to the instructions in the OIE manual [24]. To summarize, 6-week-old SPF chickens (n = 8) and 2-week-old Cherry Valley ducks (n = 10) were inoculated via the intravenous route with 0.2 ml of a 1:10 dilution of bacteria-free allantoic fluid containing 107.8 50% egg infective doses (EID50) of the A/Chicken/Korea/IS/06 virus.
Pathogenicity and transmission studies with three avian species: To study the viral pathogenesis in three avian species, 6-week-old white Leghorn chickens (n = 8), 2-week-old Cherry Valley ducks (n = 20), and 20-week-old Japanese quails (n = 13) were inoculated via the intranasal route with 106.5 EID50 of A/Chicken/Korea/IS/06. Four hours later, four uninfected chickens and quails were placed in the isolators containing the inoculated birds. These uninfected animls were referred to as the contact group. All the birds were monitored on a daily basis for clinical signs and oropharyngeal and cloacal swab samples were collected on 2, 3, 4, 6, 7 and 10 DPI. Tissue samples were collected for virus isolation from intranasally inoculated chickens (n = 8) and quails (n = 8) that died, presumably from their infections, on 3 and 4-5 DPI, respectively. Tissue samples were also collected from ten clinically normal ducks that were euthanized on 4 DPI.
Virus titration: For virus isolation, oropharyngeal and cloacal swabs as well as tissue samples from the lung, brain, kidney and heart were homogenized in sterile PBS with antibiotics. The homogenates were titrated in CEF cells to determine the TCID50. Virus titers were calculated by the Reed and Muench method [29]. For statistical analyses, we employed the two-tailed Student's t-test. Differences between the groups were considered to be statistically significant if the comparison yielded a p value of < 0.05.
Histopathological and immunohistochemical analyses
Tissue samples collected at necropsy from the intranasally inoculated chickens, quails, and ducks were fixed in 10% neutral buffered formalin solution for 24 to 48 h and routinely processed. The samples were then embedded in paraffin for histopathological and immunohistochemical examinations. For the histopathological examination, 1.5-micrometer sections were stained with hematoxylin and eosin. Duplicate paraffin-embedded sections were stained immunohistochemically by the avidin biotin peroxidase complex method employing a mouse anti-influenza nucleoprotein monoclonal antibody (MCA400; Serotec, UK) as the primary antibody. All reactions were carried out using an automated immunohistochemistry processor (NexES IHC instrument; Ventana Medical Systems, Australia) which was previously described.
Serological tests
Sera were collected from the ducks on 4, 7, 10, 14, 17, 21, 24 and 28 DPI and their avian influenza antibody titers were determined by performing the hemagglutination inhibition (HI), agar-gel immuno diffusion (AGID) and competitive enzyme-linked immunosorbent assay (C-ELISA) tests. The HI and AGID tests were performed as described in the OIE manual [24]. The antigen used in the HI test was 4 HAU (hemagglutination units) of inactivated A/chicken/Korea/IS/06 (H5N1) virus. The commercially-available C-ELISA (AniGen AIV Ab ELISA; Animal Genetics, Korea) was performed according to the manufacturer's instructions.
Results
Pathogenicity and transmission of A/chicken/Korea/IS/06 in chickens, ducks and quails
As the Korean isolates from the 2006/2007 outbreak had almost identical genome sequences, animal tests were performed with A/chicken/Korea/IS/06, which was isolated from the index chicken case in Korea. The intravenous inoculation of the chickens and ducks with 107.8 EID50 of the virus induced 100% mortality in chickens and 50% mortality in ducks with mean death times (MDTs) of 1.0 and 4.0 days, respectively. The intranasal inoculation with 106.5 EID50 of the virus also caused 100% mortality in chickens and quails but ducks showed no mortality, although they did exhibit mild clinical signs of slight depression and an unusual head tilt during the observation period. The MDTs in the chickens and quails were 3.0 and 4.8 days, respectively (Table 1).
To investigate the transmissibility of A/chicken/Korea/IS/06 in chickens and quails, uninoculated animals were placed with the intranasally inoculated groups four hours after inoculation. These contact chickens and quails showed 100% mortality with MDTs of 5.3 and 7.5 days, respectively (Table 1).
Virus shedding
We measured the virus titers in cloacal and oropharyngeal swabs taken from the intranasally inoculated birds on 2, 3, 4, 6, 7, and 10 DPI. All three avian species shed virus into their oropharynx and cloaca and the virus was already detected on the first day of testing (2 or 3 DPI). The chickens and quails shed virus right up until they died. The quails shed virus up to 6 days as opposed to 3 days for chickens (Table 2). Although the ducks did not die from their infections, they also shed virus on 2, 3, and 4 DPI, mainly via the oropharyngeal route. Thereafter, the ducks stopped shedding the virus. In all three species, the mean titers that were shed in the oropharynx were higher than those shed in the cloaca. This suggests that this virus mainly replicates in the respiratory tract and is more likely to be transmitted through direct contact than through the fecal-oral route. Since quails exhibited longer survival periods than chickens, we compared the shed virus titers of ducks with those of quails. The quails shed significantly higher oropharyngeal and cloacal virus titers during the virus shedding period than ducks. These observations together suggest that quails and ducks present a significant problem in terms of controlling HPAIV transmission; quails shed high virus titers over a relatively longer period before dying while ducks were asymptomatic and shed virus for 4 days after infection.
Virus distribution in tissues
To investigate the distribution of the virus in the intranasally infected chickens, ducks and quails, we collected samples of the lung, brain, kidney and heart from the chickens and quails that died on 3 and 4-5 DPI, respectively, and from ducks that were euthanized on 4 DPI. All examined tissues contained the virus (Table 3). However, the virus titers in the duck tissues were significantly lower (2.5 to 4.5 log10 TCID50/g) than the virus titers in the corresponding chicken (4.9 to 6.4 log10 TCID50/g) and quail (5.8 to 7.1 log10 TCID50/g) tissues. The virus titers in the chicken and quail tissues did not differ significantly except in the brain: the quails had significantly higher virus titers in the brain than chickens (p < 0.01, Table 3).
Serological analysis
Sera were collected from the intranasally inoculated ducks on 3, 7, 10, 14, 17, 21, 24 and 28 DPI. The infected chickens and quails were not tested because they did not present sero-conversion given their early death. It was revealed that a serological response was detected in ducks as early as 4 DPI by the AGID and C-ELISA tests (4 out of 11, 36.3% and 8 out of 10, 80% respectively) and 7 DPI by the HI test (1.5 ± 1.6 log2 HI titer) (Fig. 1). The antibody titers in the ducks were maintained for the duration of the experimental period, which ended 28 DPI.
Pathological features
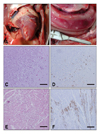
The major clinical signs of the virus-infected chickens were cyanosis, edematous combs and wattles, depression and death. The quails showed depression before death while the ducks showed only mild respiratory signs and slight depression. Upon postmortem examination of the chickens that died from the infection and the ducks that were euthanized on 4 DPI, the predominant lesions in both animals were multifocal, partly coalescent, with hemorrhagic necrosis of the pancreas (6 out of 8 chickens, 5 out of 8 ducks). Petechial hemorrhage of the cardiac fat pad was also observed in 5 out of 8 chickens (Fig. 2). Only slight gross lesions were observed in the infected quails. Histopathologically, necrosis and inflammation were observed in multiple organs of the chickens, whereas moderate meningoencephalitis including perivascular cuffing, severe nonsupprative necrotizing myocarditis and pancreatic epithelial necrosis and vacuolation were observed in the ducks.
Upon immunohistochemical analysis, viral antigens were mainly detected in the parenchymal cells of multiple chicken organs except for the intestinal tract. In the duck tissues, the virus was detected in encephalic neuronal cells, glial cells, purkinje cells, the cardiac muscle and alveolar macrophages (Fig. 2). The histopathological and immunohistochemical lesions in the quail tissues were similar to those in the chicken tissues (data not shown).
Discussion
The introduction and spread of the H5N1 HPAIV involved multiple viruses whose haemagglutinin genes were genetically related to that of the A/goose/guangdong/96 lineage virus from Southeast Asia [10]. Since their evolution, these viruses have been continuously changing, both antigenically and genetically whilst the range of their hosts have expanded to include humans [4,5,11,14,16,21,32,35,42]. As a result, the H5N1 HPAIV poses a significant and direct threat to global human health, which is of great concern [7,9,27,31,42].
Korea experienced two outbreaks of H5N1 HPAIV in the winter seasons of 2003/2004 and 2006/2007. Previous genetic analyses have shown that the Korean H5N1 HPAIVs isolated in 2003/2004 and 2006/2007 are both genetically linked to the A/goose/guandong/96 lineage [18,19]. However, it has also been shown that the 2006/2007 H5N1 HPAIV had a closer relationship to the A/bar-headed goose/Qinghai/5/2005-like lineage rather than the A/chicken/Korea/ES/2003 virus isolated in 2003/2004 in Korea [19]. To determine whether these genetic changes are accompanied with biological changes, we investigated the viral characteristics of a H5N1 HPAIV that was isolated in 2006/2007 by the experimental infection in three avian species (chickens, ducks and quails).
It has previously been shown that avian H5N1 viruses have different pathogenic potentials in avian species and mammals that range from the complete absence of clinical disease to severe neurological dysfunction and death [2-4,14,15,18,23,28,35,36,41,45,48]. Our experiment showed that the A/chicken/Korea/IS/06 virus replicated readily in the three species examined and could be transmitted to susceptible contact birds. However, the viral titers in ducks which showed systemic infection were low compared with those in chickens and quails. In addition, the ducks showed no mortality after being infected via the natural infection route. Previous reports have suggested that quails play a role in the host adaptation of H5N1 viruses that allow them to be transmitted from aquatic reservoir birds to other avian species [33]. Our results showed that quails shed similarly high amounts of the virus as chickens but for longer periods (up to 6 days before death) while ducks shed the virus without any clinical signs. This suggests that both quails and ducks may play important roles in H5N1 HPAIV transmission. We requested information regarding the pathogenicity of the A/chicken/Korea/IS/06 virus in mammalian species from the Centers for Disease Control and Prevention (USA) and received a report on the pathogenicity of this virus in mice and ferrets. This report revealed that the virus had features that were generally consistent with a high pathogenetic phenotype in mammals (data not shown). In contrast, the A/chicken/Korea/ES/2003 virus was reported to show low pathogeneticity in mammals [18]. These observations are consistent with previous reports showing that the H5N1 HPAIV genetically linked to the A/goose/guangdong/96 lineage varied significantly in their pathogenicity in different host species [2-4,14,18,28,41,45].
In the past, influenza viruses in aquatic birds were found to preferentially replicate in the gastrointestinal tract, usually without producing clinical signs, and to be mainly transmitted via the fecal-oral route [13,43]. However, the biology of the H5N1 influenza virus in waterfowl appear to be changing because most highly pathogenic H5N1 viruses that have been isolated from dead wild birds since late 2002 are excreted at high levels in the trachea (upper respiratory tract) rather than in the cloaca [2,14,26,36]. We also observed similar results when we experimentally inoculated three avian species with A/chicken/Korea/IS/06, as the virus titers in the oropharyngeal swabs of the inoculated birds were much higher than those in the cloacal swabs. These observations suggest that the transmission route of avian influenza viruses has shifted from the fecal-oral route to the oral-oral route or via some other route [1,36].
As ducks can be infected with HPAIV without exhibiting any clinical signs, active surveillance will be needed to detect HPAIV infections. Serological tests such as HI, AGID and C-ELISA tests have been used to detect the antibodies of avian influenza viruses [30,34,47]. The C-ELISA system is more sensitive and specific than the AGID test and as sensitive and specific as the HI test [47]. Therefore the C-ELISA has been established for a rapid serological diagnosis, independent of infected animal species [34]. In our experiment, although there was a high degree of correlation between C-ELISA and AGID for group specific antibody detection during the experimental period, the intensity of the precipitation band in agar-gel was obscured after 17 DPI (data not shown). Due to the fact that the AGID method may not be sensitive enough to detect low levels of viral antibody, C-ELISA can be a more useful method for the detection of antibodies in low levels at later periods of infection. Notably, our study is the first to show how ducks respond serologically to H5N1 HPAIV infection over time. However, it should be noted that these serological responses may vary depending on the inoculated virus and various host species factors such as the age of the host and its susceptibility to virus-induced diseases [2,17,25].
In summary, we have characterized a H5N1 virus which was isolated in 2006 in Korea. We have shown that the pathogenicity of the virus varies significantly in different host species and that it is mainly transmitted via the oral to oral route. We also investigated the duration of the infective stage in the three host species examined and the immune response generated by the asymptomatic duck host species. Our observations suggest that we must closely monitor duck and quail farms for the early detection of H5N1 HPAIV, thereby preventing further transmission to other avian or mammalian hosts [46].



 XML Download
XML Download