

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Taxol is a well-known anticancer drug used for many types of cancers, including breast, ovarian, and non-small cell lung cancers [4,22]. Taxol is purified from Taxus brevifolia and acts as a microtubule-targeting anticancer drug by hindering the depolymerization of microtubules within cancer cells [21]. The effects of taxol on a variety of immune cells have been studied extensively. In taxol-treated macrophages, the expression levels of inducible nitric oxide synthase were elevated and the production of interleukin-12 (IL-12), which is a critical cytokine in innate and cell-mediated immunity, was increased [10,14]. Furthermore, it was suggested that taxol might enhance the cytotoxic activity of natural killer cells [12]. Dendritic cells (DCs), the specialized antigen-presenting cells that prime naïve lymphocytes for host immune responses, are a likely target of taxol [1]. However, the effects of taxol on DCs have not been fully elucidated.
Many anticancer drugs destroy not only cancer cells, but also immune-related cells and bone marrow cells. The destruction of these latter cells results in immunosuppression and failure of hematopoietic homeostasis [19]. Interestingly, our previous study demonstrated that taxol induced the altered maturation of DCs by the enhancement of surface maturation markers, a low percentage of apoptotic cells, and a low proliferation of allogeneic splenocytes [6]. This study investigated the mechanism by which taxol induces DC survival and demonstrated that taxol sustained DC viability by protecting against cytokine withdrawal-induced apoptosis.
Materials and Methods
Animal and reagents
C57BL/6 (H-2Kb) and BALB/c (H-2Kd) mice were purchased from Orient BIO (Korea) and maintained in the animal facility of our laboratory. Female mice (7~12 week of age) were used in this study. All animal experiments were performed based on the guideline of Jeju National University for laboratory animal use and care. Taxol (Sigma, USA) purified from Taxus brevifolia was dissolved in dimethyl sulfoxide (Sigma, USA).
Generation of DCs
DCs were cultured as described previously [6]. In brief, bone marrow cells were harvested from 7~12-week-old C57BL/6 mice [8] and cultured in 6-well culture plates using RPMI 1640 media (Invitrogen, USA) containing 5% fetal bovine serum (Invitrogen, USA), 2 mM L-glutamine, 100 U/ml penicillin/streptomycin (Invitrogen, USA) and 10 ng/ml granulocyte-macrophage colony-stimulating factor (GM-CSF; Biosource International, USA). The floating cells were used as DCs 6~10 days after culture. DCs generally consisted of > 85% CD11c+ cells, as measured by flow cytometric analysis.
Quantitation of DC viability
To measure the viability of DCs, we analyzed the DC population using cell size-based flow cytometric analysis. The population of viable DCs in the dot plot was gated and confirmed based on the expression of surface DC markers, CD11c and major histocompatibility complex (MHC) class II molecules. For this, phycoerythrin-labeled anti-CD11c antibody and fluorescein isothiocyanate (FITC)-labeled anti-MHC class II antibody were used (all from BD Biosciences, USA). Cell viability was confirmed by the trypan blue exclusion test and annexin V-FITC/propidium iodide (PI) staining (Biosource International, USA).
Measurement of cytokine production
DCs were treated in 6-well culture plates with medium alone or with 1 or 5 µg/ml taxol for 24 or 48 h. The supernatants were harvested from the cultures and used for the determination of IL-12 and tumor necrosis factor-α (TNF-α) production, both of which are important cytokines for DC function [2]. Cytokine concentrations were measured by using CytoSet antibody pairs (Biosource International, USA) by enzyme-linked immunosorbent assay (ELISA) according to the manufacturer's instructions.
Flow cytometric analysis
DCs were stained for flow cytometric analysis as described previously [8]. Annexin V-FITC/PI staining was performed according to the manufacturer's instruction. Stained cells were analyzed using FACSCalibur with CellQuest software (Beckton Dickinson, USA).
Western blot analysis
Western blot analysis was performed as described in a previous study [7]. In brief, DCs were treated in 6-well culture plates with 5 µg/ml taxol for 6, 24 or 48 h. DC lysates were harvested and the protein concentrations were determined using Bradford protein assay (Bio-Rad, USA). Proteins were separated in a 12% polyacrylamide gel and blotted onto nitrocellulose membranes. Anti-Bcl-2, anti-Bcl-xL, and anti-Bax antibodies (Santa Cruz Biotechnology, USA) were used, followed by their respective horseradish peroxidase-labeled secondary antibodies (Santa Cruz Biotechnology, USA), to detect specific proteins. As an internal control, anti-β-actin antibody (Sigma, USA), followed by its respective secondary antibody (Santa Cruz Biotechnology, USA), was used to detect β-actin. Protein bands were visualized using SuperSignal West Pico Chemiluminescent Substrate (Pierce Biotechnology, USA).
Confocal microscopic analysis and inhibitor treatment
DCs were treated on chamber slides with medium alone, 5 µg/ml taxol for 3 h. In brief, cells were fixed with 2% paraformaldehyde-containing phosphate-buffered saline and then stained with 4 µg/ml tetramethylrhodamine isothiocyanate-labeled anti-nuclear factor-κB (NF-κB) p65 monoclonal antibody (Santa Cruz Biotechnology, USA). The pictures of stained DCs on chamber slides were obtained using a confocal microscope (Olympus Optical, Japan). For the NF-κB inhibitor assay, DCs were treated with N-p-tosyl-L-phenylalanine chloromethyl ketone (TPCK; Sigma, USA).
Results
Taxol increases DC viability
In the flow cytometric analysis, two different cell populations were gated based on cell size; region R1 included DCs, as confirmed by staining for cell-specific surface markers, CD11c and MHC class II, whereas region R2 contained contaminating cells, mainly dead cells (Fig. 1). In preliminary experiments, we confirmed that the viable DCs show higher FSC/SSC, which means bigger than dead DCs since the cell size of DCs are reduced upon cell death. Based on this analysis, the taxol-treated DCs (TaxolDCs) showed significantly higher viability than medium alone-treated DCs (ContDCs). These results suggest that taxol may protect DCs from cytokinewithdrawal-induced cell death.
The enhanced cytokine production of DCs treated by taxol
The TaxolDCs treated with 5 µg/ml of taxol produced higher amounts of both cytokines than the ContDCs, at 24, 48 h of incubation time (Fig. 2). However, the taxol concentration used was critical for the level of cytokine production; 1 µg/ml of taxol induced only marginal production of these cytokines. These results suggest that taxol enhances the production of cytokines that are critical for cell-mediated and innate immunity.
Taxol increases Bcl-xL expression, an anti-apoptotic protein in DCs
Western blot analysis showed that Bcl-xL expression in TaxolDCs was increased after 6 h of treatment, whereas the expression levels of Bcl-2 and Bax were not increased (Fig. 3). The protein expression levels were calculated in comparison to the level of β-actin (internal control). The results suggest that taxol enhances DC viability via the increase of the anti-apoptotic protein Bcl-xL as a potential mechanism of action.
NF-κB signaling in taxol-induced DC survival
Using confocal microscopy, the mobilization of NF-κB p65 subunit molecules from the cytosol to the nucleus were much greater in TaxolDCs in comparison to ContDCs (Fig. 4A). Furthermore, TPCK, which is an inhibitor of the NF-κB pathway, significantly decreased the viability of TaxolDCs, but not that of ContDCs (Fig. 4B). These results suggest that taxol sustains DC survival via the NF-κB pathway.
Discussion
Our previous study demonstrated that taxol induces the altered maturation of DCs and increased their viability in comparison to the medium alone [6]. However, the mechanisms by which taxol enhances DC survival have not been elucidated. The present study confirmed taxol's enhancement of DC viability using different assays than those used in the previous study. Taxol-treated DCs appeared as active effector cells, producing major cytokines involved in immune responses. Importantly, taxol increased the expression of the anti-apoptotic protein Bcl-xL, and increased the mobilization of p65 subunit molecules into the DC nucleus. Furthermore, we found that the inhibition of NF-κB by TPCK decreased DC viability.
The Bcl-2 family proteins have been known to act as apoptosis-related proteins in immune cells including DCs [5,15,16]. Among these proteins, Bcl-2 and Bcl-xL show anti-apoptotic activity, whereas Bax has pro-apoptotic activity. The present study demonstrated that the expression of Bcl-xL increased in DCs at early time-points of taxol treatment, whereas that of Bcl-2 was not increased. In a previous study, transduced Bcl-xL in DCs showed anti-apoptotic activity in a mouse tumor model [18]. Therefore, it seems likely that Bcl-xL was at least partially responsible for enhancing DC viability. As a future study, the knockdown study of Bcl-xL using siRNA may further confirm their role of taxol-treated DCs. Although the expression of Bax was increased in TaxolDCs, the expression of Bax occurred later than that of Bcl-xL, which implies that the pro-apoptotic effects of Bax might be minimized in the DCs during the treatment of taxol, 48 h.
NF-κB is closely involved in the survival and maturation of DCs [11,17,20]. To investigate further the mechanisms of taxol-induced changes, the viablity of TPCK, an inhibitor of the serine protease that inhibit NF-κB signaling pathway in cells [20], pretreated DCs were measured. It was observed that TPCK significantly decreased the viability of TaxolDCs, but not that of ContDCs (p < 0.05). In our previous study, the optimal concentration of TPCK (4 µM) that could inhibit NF-κB signaling pathways without affecting the survival of ContDCs was determined [9]. These results suggest that NF-κB is involved in the survival of TaxolDCs.
Immunotherapy using DCs alone can provide cancer patients with strong benefits, including the eradication of cancer cells located in difficult sites to be reached via anti-tumor immunity [13]. However, the lack of tumor-specific antigens may hamper the development of a complete cure for cancers, although DCs have a strong ability to generate antigen-specific cytotoxic T lymphocytes [1]. Thus, recent experimental approaches to immunotherapy have focused on the application of DCs in combination with chemotherapy, specifically taxol [3,23]. This study has demonstrated that a popular anticancer drug, taxol, enhances DC viability. These findings provide a basic rationale for the use of taxol in DC immunotherapy and insights into the development of combinational DC therapies with other anticancer drugs that do not harm the viability or activity of DCs.

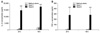


 XML Download
XML Download