

 PDF
PDF Citation
Citation Print
Print



Abbreviations
AID
activation-induced deaminase
BCR
B cell receptor
GC
germinal center
ICOS
inducible T cell costimulatory
ICOSL
inducible T cell costimulatory ligand
IFN
interferon
iNKT
invariant natural killer T
LPS
lipopolysaccharide
MHC
major histocompatibility complex
MZ
marginal zone
NKT
natural killer T
SLAM
signaling lymphocytic activation molecule
TCR
T cell receptor
TD
T cell-dependent
Tfh
follicular helper T
Th
T helper
TI
T cell-independent
TLR
toll-like receptor
INTRODUCTION
The activation of Ag-specific B cells via B cell receptor (BCR) is an essential requirement of the Ab response. However, a sufficient Ab response requires not only BCR-mediated signaling but also participation of CD4+ T cells or activation of innate immune receptors on B cells (123). Since protein Ag-specific B cells usually require major histocompatibility complex (MHC) class II-restricted Ag presentation and CD4+ T cell help, this kind of Ab response is called as T cell-dependent (TD) Ab response (4). On the other hand, Ab responses to sugar or lipid Ags can occur in the absence of CD4+ T cells as a T cell-independent (TI) Ab response since those Ags with repetitive epitopes engage multiple BCRs or are frequently associated with pathogen-associated molecular patterns (5). However, the TI Ab responses can be boosted by T cells (6). These T cell-promoted TI responses make it difficult to clearly distinguish between the TD and TI Ab responses. In this review, we would like to discuss the participation of innate or innate-like T cells in the TI Ab responses and the interaction between innate-like T cells and B cells.
INNATE OR INNATE-LIKE LYMPHOCYTES
Innate T and B lymphocytes have recently attracted much attention. Although different opinions exist regarding innate lymphocytes, in this review, we refer to innate lymphocytes as cells having T cell receptors (TCRs) or BCRs, thereby distinguishing innate lymphoid cells from innate lymphocytes (7). Differently from conventional lymphocytes, innate lymphocytes obtain a memory phenotype, such as rapid cytokine secretion or rapid Ab production, during development in the absence of previous Ag exposure. Their rapid and strong responses support the initial priming of conventional naïve T cells, as exemplified in natural killer T (NKT) cells that bridge innate and adaptive immunity by secreting interferon (IFN)-γ in response to IL-12 produced by dendritic cells (8). As bona fide conventional memory lymphocytes are absent at the time of birth, innate or innate-like lymphocytes must function as first-line defenders in neonates and possibly in adults as well (7). γδ T, NKT, and B-1a cells are well-established innate lymphocytes that function as both effectors and regulators of immunity (39).
B-1 and marginal zone (MZ) B cells – 2 distinct types of innate or innate-like B cells
B-1 and MZ B cells participate cooperatively in the TI Ab response to blood-borne bacteria (10), but they function via different mechanisms. B-1a cells, which are CD5+ B-1 cells derived from fetal hematopoietic stem cells and are localized in serosal cavities, are mostly autoreactive and also cross-reactive to common pathogens (11). The stimuli for functional activation of B-1a cells are usually through pattern recognition receptors, such as toll-like receptor (TLR)-4, since BCRs of B-1a cells are chronically activated by autoantigens (1213). Stimulation of B-1a cells with lipopolysaccharide (LPS) leads to their migration out of the serosal cavities into the spleen and consequent secretion of natural Abs and granulocyte macrophage colony-stimulating factor (1415). B-1a cells readily migrate into the lung, intestine, or regional lymph nodes upon infection of these organs (1617). Natural IgM Ab secretion by B-1a cells has been found to be constitutive and dependent on Blimp-1, but Blimp-1-independent IgM Ab secretion by B-1a cells has also been noted, suggesting that B-1a cells do not exist as a single population (18). The subdivision of B-1a cells into homeostatic and induced responding populations has been suggested (19).
In contrast, MZ B cells are pre-activated B cells that are localized around the splenic marginal sinus. They can rapidly respond to blood-borne pathogens that are trapped by MZ macrophages around the sinus (2). The development of MZ B cells is important for proper responses against blood-borne pathogens. The appropriate selection of BCR specificities against common pathogens is essential for MZ B cell responses because the selection of autoreactive MZ B cells can be potentially harmful to the host (20). MZ B cells develop from bone marrow precursor cells through the stages of splenic transitional B and MZ precursor cells (21). The developmental decision from transitional B cells to MZ B or follicular B cells is determined by BCR specificities and Notch signaling. Although MZ B cells have some stem cell-like properties to repopulate themselves, new MZ B cells can develop from transitional B and MZ precursor cells (22).
In summary, B-1 and MZ B cells are the two main populations of B cells responsible for TI Ab responses, but they are regulated by different mechanisms. Here we would like to discuss the roles of innate-like T cells in TI Ab responses by B-1 and MZ B cells.
TD Ab RESPONSE
The ultimate outcome of the classical TD Ab responses is the generation of high-affinity Abs via the interaction between germinal center (GC) B cells and follicular helper T (Tfh) cells. It is clear that Tfh cell differentiation is crucial for the GC reaction leading to the selection of high-affinity Ab responses against protein Ags (4). For the GC reaction, activated B and conventional CD4+ T cells interact closely in cognate or non-cognate manners, resulting in their respective differentiation into GC B and Tfh cells through Bcl-6 upregulation (23). Although the CD40–CD40L interaction is essential for the GC reaction, the GC reaction requires more T cell–B cell interactions via inducible T cell costimulatory (ICOS)–inducible T cell costimulatory ligand (ICOSL) and signaling lymphocytic activation molecule (SLAM)–SLAM interactions (24). Many T cell–B cell interactions fail to progress to the GC reaction, instead resulting in extrafollicular Ab responses (25). Targeted ablation of Tfh cells via CD4+ T cell-specific deletion of Bcl-6 could not inhibit Ab responses as effector CD4+ T cells, such as T helper (Th) 1 and Th2 cells, could promote extrafollicular TD Ab responses (2627). Extrafollicular TD Ab responses lead to the generation of Ag-specific B cells with a low level of somatic hypermutation, but the affinity constants of Abs generated from these responses are far lower than those of Abs generated by the Tfh cell-dependent GC reaction and affinity maturation (27). Therefore, the TD Ab responses can occur with or without the GC reaction and the Tfh cell differentiation.
TI Ab RESPONSE
TI Ab responses are conventionally divided into type-1 (TI-1) and type-2 (TI-2) responses. Whereas TI-1 Ab responses require the engagement of pattern recognition receptors, such as TLR-4 or TLR-9, in addition to BCR engagement, TI-2 responses require the engagement of at least 10 molecules of BCRs by highly repetitive Ags (28). The fact that TI Ab responses can occur in the absence of T cells does not mean that T cells do not play a role in these responses. The involvement of T cells in TI Ab responses, especially TI-2 responses, has been well established (629). Furthermore, T cell involvement in TI Ab responses is accompanied by class switching and a low level of somatic hypermutation, which are dependent on the expression of activation-induced deaminase (AID) (30). In fact, AID expression is upregulated during extrafollicular TD and TI Ab responses and is thus not unique to the GC reaction (25). Interestingly, B-1a cells are known to upregulate AID expression upon LPS stimulation (3132). The identities of T cell subsets in the TI Ab responses are discussed in detail in the subsequent sections.
Because infectious agents contain both TD and TI Ags, both TD and TI Ab responses proceed concurrently upon infection in most cases. Thus, TD Ag-specific B cells can be initially activated by innate stimuli or the complement pathway and proliferate in a TI manner, thereby blurring the distinction between TD and TI Ab responses (33). Considering the extremely rare chance of cognate interactions between conventional Ag-specific B and T cells, initial TI B cell responses may facilitate TD Ab responses and enhance the possibility of cognate interactions between rare Ag-specific B and T cells via the expansion of Ag-specific B cells (1). Interestingly, early IgM secretion by B-1a cells or initially stimulated B cells can enhance the overall Ab response via promotion of Ag presentation by dendritic cells (34) or positive feedback through FcμR (35). Therefore, TD Ab responses can be supported by early TI Ab responses that can be actually supported by T cells.
INVOLVEMENT OF INNATE OR INNATE-LIKE T CELLS IN TI Ab RESPONSE
NKT cells in the MZ B cell immune response
NKT cells have direct and indirect effects on B cell Ab responses. First, NKT cells amplify inflammatory or cytokine responses. They rapidly secrete IL-4 or IFN-γ in response to IL-12 produced by dendritic cells during early infection and facilitate adaptive immune responses of conventional T and B cells (836). α-Galactosylceramide, an NKT cell agonist presented within CD1d, can be used as an adjuvant the TD Ab responses (37). Notably, NKT cells can form a cognate interaction with CD1d-expressing B cells. In particular, MZ B cells more highly express CD1d than other subsets of B cells, making them excellent Ag-presenting cells for NKT cells (38).
When a given B cell is specific to a lipid Ag that is presented within CD1d, an NKT cell can provide cognate help to the B cell (3940). The CD1d-restricted cognate interaction between NKT and B cells leads to the development Bcl-6-expressing follicular helper NKT cells, but interestingly, long-lived plasma cells are not generated by this CD1d-mediated cognate interaction (41). This response has been described as the TD type 2 (TD-2) Ab response because this is different from the classical TD response and usually fails to generate high-affinity Ag-specific B cells (42).
When a given B cell specific to a protein Ag is activated by the specific Ag in combination with innate stimuli, such as LPS, the activated B cell can present endogenously generated NKT cell ligands as well as peptides (43). This suggests that NKT cells have the potential to support B cells with different types of BCR specificities in both TD and TI Ab responses independently of the cognate interaction via MHC class II. It should be noted that NKT cells can provide non-cognate help for both MZ B cell TI and follicular B cell TD Ab responses because NKT cells are recruited to the regional lymph nodes under inflammatory conditions and provide IL-4 (3644). For high-affinity Ab TD responses, an exact MHC class II-mediated cognate interaction is crucial, and the CD1d-mediated cognate interaction cannot replace the requirement of the MHC class II-mediated cognate interaction. In summary, NKT cells can support the CD1d-dependent TI Ab response via a cognate NKT-B cell interaction or in a non-cognate manner and boost the classical TD Ab response by supplying cytokines and costimulatory molecules.
γδ T cells and autoreactive GCs
The GC reaction can be induced by γδ T cells in αβ T cell-deficient mice (4546). γδ T cells have been found in some GCs of humans, sheep, and TCR α−/− mice (47). The Vγ9Vδ2 T cells, the most common human γδ T cells in human peripheral blood, are activated by butyrophilin 3A1 that binds to phosphorylated metabolites derived from microbes and transformed cells (48) and secrete inflammatory cytokines and enter into GCs themselves, becoming γδ Tfh cells (49). However, γδ T cell-supported GCs are fewer than those in the classical TD Ab response. The GC formation is reduced in athymic nu/nu mice, but recovers to normal levels after adoptive transfer of conventional αβ T cells (50). Remarkably, this γδ T cell-driven GC response was induced by repeated parasitic infections and resulted in an enhancement of autoreactive B cells instead of pathogen-specific B cells (51). This GC reaction appears to be unique in that the γδ T cells provide help for autoreactive B cells in a non-cognate fashion. This implicates the importance of γδ T cells in the pathogenesis of autoimmune diseases such as systemic lupus erythematosus and B cell dysfunction in acquired immune deficiency, as spontaneously developed GCs harbor autoreactive B cells with somatic hypermutations (52). At present, the cellular and molecular mechanisms underlying this interaction are not well understood. It would be interesting to address the identities of B cell-helping γδ T cells and whether innate B cells are involved in the collaboration with γδ T cells.
B-1 helper T cells in B-1a cell immune response
B-1 cells are divided into CD5+CD11b+ B-1a and CD5−CD11b+ B-1b cell types, which develop from fetal and adult hematopoietic stem cells, respectively (11). B-1a cells are thought to produce natural Abs in a TI manner, as innate stimuli or cytokines, such as IL-5, induce Ab production (53). Many carbohydrate and lipid Ags are believed to be recognized by B-1a cells, as noted in a report on B-1a cells expressing receptors for blood group A carbohydrates (54). Several B-1b cell Ags have been reported (55), and reportedly, B-1b cells form a TI memory against Borrelia hermsii (56). The involvement of T cells in B-1 cell Ab responses is not well investigated, but an active interaction between B-1 and CD4+ T cells is plausible because B-1 cells are excellent Ag-presenting cells for T cells (57). The combination of IL-4, IL-5, and the CD40–CD40L interaction was suggested to be a mechanism underlying CD4+ T cell help for B-1a cells (58).
NKT cells were thought to be good candidates as helpers of B-1a cells, according to a previous finding that NKT cells are helpers of B cells expressing BCRs for blood group A carbohydrates (59). However, in the case of response to α (13) Gal epitopes, the requirement of conventional CD4+ T cells in addition to NKT cells was demonstrated (60). Therefore, both conventional CD4+ T and NKT cells are plausible candidate helpers for B-1 cell Ab responses. Previously, we attempted to identify B-1a cell subpopulations for effector Ab-secreting function and/or repopulation with stem cell-like property and observed that B-1a cells conjugated to CD4+ T cells were superior in terms of IgM Ab production (61). The serosal CD4+ T cells contained a unique memory phenotype T cells that expressed a high level of CD49d (integrin α4) and developed spontaneously before 2 weeks of age. Upon stimulation with phorbol myristate acetate and ionomycin, these cells rapidly secreted Th1-type cytokines, such as IFN-γ, tumor necrosis factor-α, and IL-2. The capability of these cells to provide B-1a cell help was clearly revealed in the experiments with co-adoptive transfer of B-1a cells and serosal CD49dhighCD4+ T cells into lymphocyte-deficient mice and co-culture of these two types of cells. The CD49dhighCD4+ T cells expressed high levels of integrin α4β1 and α6β1, suggesting their capability to enter peripheral inflammatory sites and migrate via interaction with laminins (62).
We assume that the serosal CD49dhighCD4+ T cells are B-1 helper T cells capable of boosting B-1 cell secretion of natural Abs. A similar CD49dhighCD4+ T cell population was also noted in humans (63). At this point, the mechanism by which these CD49dhighCD4+ T cells assist B-1a cells is unknown. Different cellular and molecular mechanisms are possible, including: 1) a bystander interaction through costimulatory molecules, such as the pairs CD40–CD40L and ICOS–ICOSL independent of TCR–MHC class II; 2) TCR recognition of idiotypic peptides derived from immunoglobulin heavy chain forming an Ab idiotype–anti-idiotype network (64); or 3) MHC class II-dependent TCR recognition of non-peptide Ags, such as glycosylated MHC class II (65). In any case, the functional mechanism of B-1 cell help by these innate-like CD4+ T cells needs to be investigated in the future.
SUGGESTION OF A NEW SUB-CLASSIFICATION OF TD Ab RESPONSES
In this review, we discussed the role of innate-like T cells in TI Ab responses, which hampers the clear distinction between TD and TI Ab responses in in vivo humoral responses. The classical TD Ab response generates high-affinity Ab through the GC reaction and requires absolute involvement of Tfh cell differentiation. However, the TD Ab response also requires preceding multiple interactions between activated B and T cells, many of which lead to extrafollicular Ab responses. Considering multiple T-B interactions, we propose to discriminate the T-B interactions leading to the GCs or extrafollicular Ab responses as TD-1 or TD-2 Ab responses, respectively (Fig. 1). We suppose that the GC-generating TD-1 response is always accompanied with the extrafollicular TD-2 responses and that some Ab responses are predominated by extrafollicular TD-2 responses without the GC reaction. Thus, our definition of the TD-2 response is different from the TD-2 Ab response that had been previously suggested to refer to NKT cell help to B cells via CD1d-mediated cognate interaction (53). The newly suggested TD-2 response is extrafollicular and can involve both conventional CD4+ T and NKT cells. To expand the concept of the TD Ab responses, we suggest a new definition of a TD-3 Ab response, in which NKT or MHC class II-restricted T cells provide help for B cells in a non-cognate manner (Table 1). The difference between TD-2 and TD-3 responses is whether BCR delivers the antigenic peptide or non-peptide Ag for the cognate interaction between TCR and MHC class II, CD1d, or other Ag-presenting molecules (Fig. 2). Therefore, critical points distinguishing TD-1, TD-2, and TD-3 responses are whether the response leads to the GC reaction (TD-1) or extrafollicular response (TD-2 and TD-3) and whether B cell presents peptide or lipid derived from the exact BCR-bound Ag to solicit T cell help (TD-1 and TD-2) or B cell is helped by bystander T cell (TD-3). The TD-3 response is less likely to occur between resting B and T cells, but occurs between activated B and T cells upon infection or other pathogenic stimuli. The non-cognate or bystander interaction between T and B cells is important for the overall Ab responses as previously reported (57666768). Another example of non-cognate B cell help via MHC class II is when CD4+ T cell recognizes idiotypic peptides from immunoglobulin heavy chain variable domain present within the MHC class II molecule (64). If NKT cell help to B cells specific for protein Ags occurs via CD1d, this CD1d-mediated interaction is non-cognate with respect to BCR specificity, and this Ab response should belong to a TD-3 response. The B cell help by γδ T cells is regarded as a non-cognate TD-3 response, but appears to be exceptional in that this response can produce autoreactive GCs (4751). It is interesting to address whether innate-like serosal CD49dhighCD4+ T cells provide B-1a cells in a non-cognate and MHC class II-dependent manner.
Figure 1
The sub-classification of the TD Ab responses. In the left panel (depicted as TD-3 response), a B cell is activated by an innate stimulus (not depicted, a TI response) and/or an innate-like T cell through non-cognate interaction involving TCRs and Ag-presenting molecules, such as MHC or CD1d (not depicted), irrespective of the BCR specificity (TD-3 response). The activated B cell can proliferate and advance further to interact with other T cells, eventually resulting in a cognate interaction involving TCRs and peptides derived from BCR-bound proteins. This interaction may lead to an extrafollicular Ab response (TD-2 response, right upper) or eventual GC formation (TD-1 response, right lower). iNKT cell can provide cognate help for MZ B cells, leading to extrafollicular MZ B cell response as shown in right upper corner.
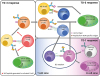
Table 1
The classification of TD Ab responses

Figure 2
Cognate and non-cognate B cell help by MHC class II- or CD1d-restricted T cells. (A) B cell specific to a protein or lipid (not depicted) Ag presents the antigenic peptide within MHC class II or lipid moiety within CD1d (not depicted) and is helped by activated T cell or NKT cell, respectively (TD-1 or TD-2 responses). (B) Even though a B cell specific to a protein Ag presents a peptide from the protein Ag within MHC class II, the B cell can solicit help from a iNKT cell via the interaction between exogenous or self-lipid Ag within CD1d and TCR (TD-3 response). This help is non-cognate with respect to BCR. (C) The non-cognate B cell help can be supported by a bystander MHC class II-restricted helper T cell, which represents another TD-3 response. This bystander helper T cell can be activated T cell specific to other peptides or innate-like T cell that acquire a memory phenotype during development. The source of peptides for bystander helper T cells may be endogenous or derived from endocytosed proteins.

CONCLUDING REMARK
In vivo B cell Ab responses are extremely complex as they involve combinations of different types of TD and TI Ab responses. In addition to the classical cognate T–B interaction, the presence of non-cognate T–B interaction and extrafollicular Ab responses blurred the clear distinction between the TD and TI Ab response. Therefore, we suggested a new classification of the TD Ab responses to incorporate gray zone TD responses in this review. Especially, the innate-like T cells are thought to be important in boosting the overall Ab responses by providing cognate and non-cognate B cell help. The importance and types of innate-like helper T cells in all types of Ab responses needs to be investigated in the future.
 XML Download
XML Download