

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Febrile seizure (FS) is defined as a seizure with fever in children younger than 6 years who have no history of epilepsy and without any obvious CNS infection, and it is the most common and unique form of seizure that only occurs during early childhood.1 Fever can be induced by various proinflammatory cytokines such as interleukin (IL)-1β, IL-6, and tumor necrosis factor (TNF)-α during infections.2 Cytokine responses to infections can influence the body temperature, and the overproduction of proinflammatory cytokines may easily overwhelm the threshold for FS in children, which might be referred to as a susceptibility to inflammation.
Generalized epilepsy with febrile seizure plus (GEFS+) is a familial epilepsy syndrome associated with FS and epilepsy that is typically inherited with autosomal dominance and variable penetrance.3 A GEFS+ family is defined if it contains more than two patients with GEFS+ phenotypes.4 FS only is the most common phenotype, constituting 44% of GEFS+ families, while so-called FS+ is the second most common phenotype (27%,) which is defined as FS at older than 6 years or in a child with both FS and afebrile generalized tonicclonic seizure.5 The genes implicated in GEFS+ are the alpha-1 neuronal voltage-gated sodium channel subunit Nav1.1 (SCN1A), the beta-1 subunit of the voltage-gated sodium channel (SCN1B), and the γ-aminobutyric acid (GABA)-A gamma-2 receptor subunit (GABRG2).4 Although the most common phenotypes in GEFS+ are fever-related seizures, whether genetic susceptibility to inflammation is implicated in GEFS+ is still unclear.
In our previous study we found elevations in both proinflammatory and anti-inflammatory cytokines such as IL-1β, IL-6, TNF-α, high mobility group box-1 (HMGB1), and IL-10 in peripheral blood after FS in children with FS but also after an afebrile seizure (AFS) in children with epilepsy.6 Moreover, IL-1β was significantly increased and was correlated with other proinflammatory cytokines including HMGB1 after an FS attack compared to febrile children without a seizure attack, implicating the cytokine network in the generation of FS.6 IL-1β has also been found to be increased in the CSF and epileptogenic brain lesions of children with epilepsy, suggesting that this cytokine is important in epilepsy initiation and progression.78
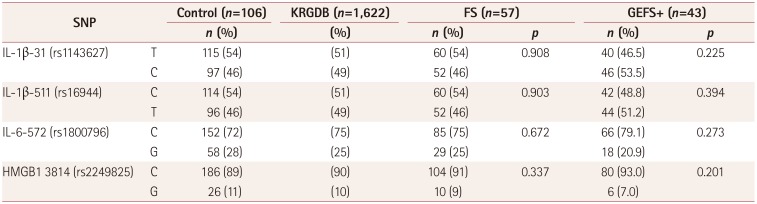
A family history of FS is reportedly present in 17–30% of children with FS.2 The influences of cytokine gene variants on the susceptibility to FS have been controversial. The IL-1β-511 promoter polymorphism was found to be associated with FS in a meta-analysis and lead to an increase in IL-1β
production,91011 but other studies have not found such associations.1213 Moreover, there have been no human studies of the correlations of actual postictal serum cytokine levels with genotypes of cytokine genetic variants in children with FS.
In this study we analyzed the correlations of postictal serum cytokine levels with the promotor variants of IL-1β, IL-6, and HMGB1 in children with FS and GEFS+.
METHODS
Patient information
Children with FS or GEFS+ who visited the emergency room of Seoul Metropolitan Government Seoul National University Borame Medical Center were enrolled in this study (Table 1). The inclusion criteria were children with FS aged between 3 months and 5 years experiencing seizures associated with fever above 38℃ and without CNS infection, neurologic deficits, or previous AFS.14 GEFS+ were diagnosed according to the criteria in the 2017 International Classification of Epileptic Syndromes15 based on the clinical history and demographic findings of patients who developed both FS and AFS with a positive family history of FS or epilepsy. We excluded FS patients whose serum CRP levels were above 1.0 mg/dL in order to avoid the influence of severe infections accompanying FS on cytokine profiles.
Blood was obtained and DNA was extracted using Qiagen DNA purification kits (Germantown, MD, USA) and frozen for genotyping and sequencing. Serum was immediately separated only from patients who visited within 2 hours after an acute seizure attack, from FS patients after an FS attack, and from GEFS+ patients after an AFS attack, and then frozen for subsequent cytokine assays. The difficulty of drawing extra blood for cytokine measurements within 2 hours after seizures in addition to the routine blood work including blood cultures meant that the number of acute serum samples available for cytokine measurements was smaller than the total number of FS and GEFS+ patients. Postictal serum cytokine levels were available from 38 FS patients, 10 GEFS+ patients, and 20 febrile control children.
Clinical data about the familial FS history, the previous FS attack, and the semiology and time duration of FS were gathered from the parents of the patients. The controls were age matched and did not have previous FS or epilepsy. This study was approved by the Seoul Metropolitan Government Seoul National University Borame Medical Center Institutional Review Board (approval numbers 20080918/06-2008-74/76), and informed consent was obtained from the parents of each child.
SNP selection and genotyping
Four variants located in the promotor regions of IL-1β-31 (rs1143627), IL-1β-511 (rs16944), IL-1β-1003 (rs1143625), and IL-1β-1473 (rs1143623), four variants located in the promotor regions of IL-6-174 (rs1800795), IL-6-572 (rs1800796), IL-6-597 (rs1800797), and IL-6-1426 (rs2069827), and the HMGB1 3814 (rs2249825) variant were initially selected from the HapMap human single nucleotide polymorphism (SNP) database (https://www.ncbi.nlm.nih.gov/probe/docs/projhapmap/) and the dbSNP database. However, IL-6-174 (rs1800795), IL-6-597 (rs1800797), and IL-6-1426 (rs2069827) were subsequently excluded due to having a minor allele frequency of lower than 0.05 (Supplementary Table 1 in the online-only Data Supplement). Hardy-Weinberg equilibrium exceed 0.05 in all patients. However, the Hardy-Weinberg equilibrium for controls was less than 0.05 for IL-1β-1003 (rs1143625) and IL-1β-1473 (rs1143623) (Supplementary Table 1 in the online-only Data Supplement), and so those variants were also excluded when analyzing the data.
Pairwise linkage disequilibrium (LD) of SNP markers was estimated using Haploview (version 4.0; http://www.broadinstitute.org/haploview). The default confidence interval algorithm of the Haploview program identified one haplotype block consisting of IL-1β-31 and IL-1β-511 from patient group data (Supplementary Fig. 1 in the online-only Data Supplement). Haplotypes were estimated using the HAPLOTYPE procedure in SAS/Genetics software (version 13.1; SAS Co., Cary, NC, USA).
Primer sequencing and genotyping
Probes and primers were designed with genomic sequence information. Genotyping was performed with the SNaPshot assay (Sequenom, San Diego, CA, USA). The person who analyzed the genotypes was blinded to the clinical data.
Cytokine measurement
IL-1β, IL-6, and HMGB1 serum levels were detected using ELISA according to the manufacturer's instructions (Panomics, Redwood City, CA, USA; R&D Systems, Minneapolis, MN, USA; and Shino-Test Corporation, Tokyo, Japan16; respectively), with detection limits of 0.27, 0.23, and 0.2 ng/mL, respectively.
Statistical analysis
The Hardy-Weinberg equilibrium of each variants was evaluated using genotype data from all groups for goodness-of-fit. The trend test, two-sided chi-square test with Yate's correction, and Fisher's exact test were used to analyze genotype differences between FS patients, GEFS+ patients, and controls depending on the mode of inheritance,17 based on the minor allele of each variants. We used the logistic regression test to assess the associations of each variants with FS and GEFS+. The dependence of postictal serum cytokine levels on various cytokine genotypes was analyzed using the Kruskal-Wallis test or Mann-Whitney U test with multiple comparisons. IBM SPSS Statistics (version 20; IBM Corp., Armonk, NY, USA), R (version 3.2.5; http://www.r-project.org), and GraphPad Prism (version 7.03; San Diego, CA, USA) were used to perform the tests. Mean values are quoted, and the criterion for significant differences was p<0.05 in all tests.
RESULTS
Characteristics of the subjects
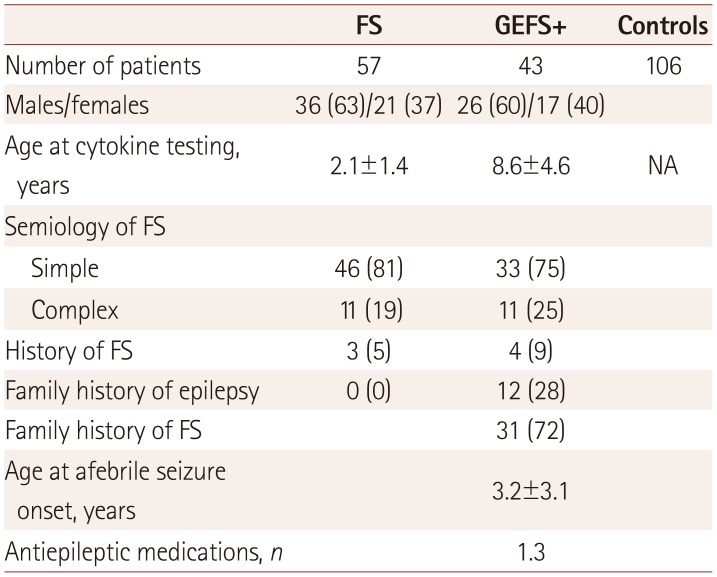
Table 1 summarizes the clinical data of the included subjects, who comprised 56 FS patients, 43 GEFS+ patients, and 108 control children. Males were more prevalent than females among the FS and GEFS+ subjects (63% and 60%, respectively). The semiology types of the FS attacks comprised 81% simple and 19% complex. Three patients had histories of febrile status epilepticus, FS sustaining more than 30 minutes.
All GEFS+ patients showed the evolution of epilepsy after an initial FS attack, and so were diagnosed by having both FS and AFS with a positive family history. Twelve patients (28%) had a family history of epilepsy and 72% of the patients had a family history of FS. The mean age at the first AFS was 3.2 years, and the GEFS+ patients had been treated with a mean of 1.3 antiepileptic medications. The mean age at testing due to an acute AFS attack was 8.6 years.
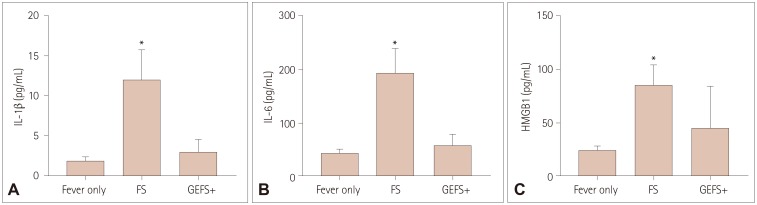
The postictal serum IL-1β, IL-6, and HMGB1 levels were significantly higher in FS patients than in controls (all p<0.05) (Fig. 1). The serum postictal cytokine levels in GEFS+ patients after AFS were higher than controls, but the difference between GEFS+ and controls showed no significance, statistically (Fig. 1).
Allele comparisons
The allele distributions in FS and GEFS+ patients are compared to controls in Table 2. We also used the Korean Reference Genome Database for controls, which contains public data for 1,622 genome data of the Korean population (http://coda.nih.go.kr/coda/KRGDB/index.jsp). The allele distributions in FS and GEFS+ patients did not differ from those in the controls.
CT genotypes of IL-1β-31 and IL-1β-511 are significantly correlated with higher postictal serum IL-1β levels in FS patients
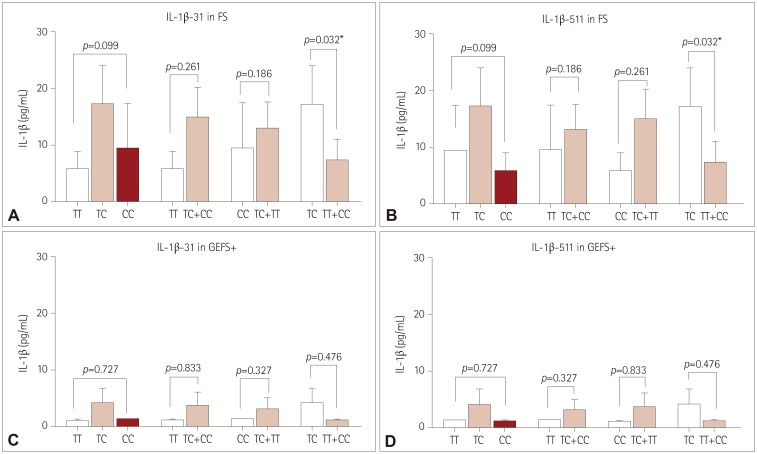
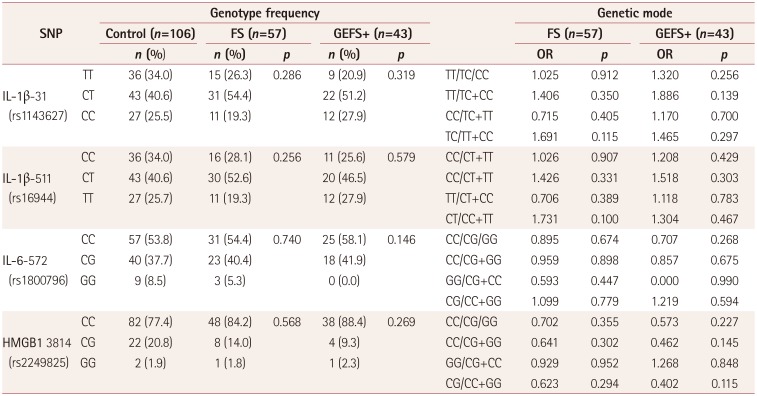
CT genotypes of IL-1β-31 and IL-1β-511 showed significant correlations with higher postictal serum IL-1β levels compared to CC+TT genotypes among FS patients (17.2 vs. 7.4 pg/mL, both p<0.05) (Fig. 2A, B). Moreover, CT genotypes of the IL-1β-31 and IL-1β-511 variants showed stronger associations with FS than did CC+TT genotypes [odds ratio (OR)=1.691 and 1.731, respectively] (Table 3).
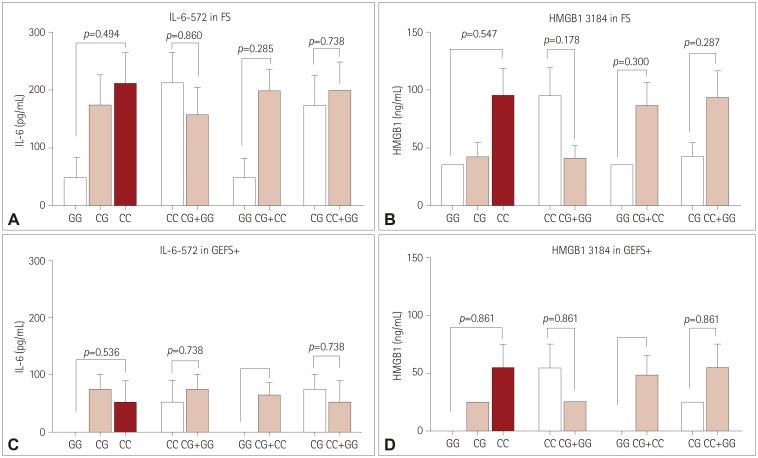
IL-6-572 and HMGB1 3814 were not associated with genotypes or correlated with postictal serum levels in FS patients compared to controls (Fig. 3A, B).
CT genotypes of IL-1β-31 and IL-1β-511 are correlated with higher postictal serum IL-1β levels in GEFS+ patients
IL-1β-31 and IL-1β-511 variants showed no statistically significant correlation with postictal serum levels in GEFS+ patients after AFS attacks. Although, CT genotypes of both IL-1β-31 and IL-1β-511 showed slightly higher postictal serum IL-1β levels compared to the CC+TT genotypes (4.1 vs. 1.2 pg/mL, both p=0.476) (Fig. 2C, D). TT genotypes of IL-1β-31 and CC genotypes of IL-1β-511 showed slightly stronger associations with GEFS+ (OR=1.886, OR=1.518; Table 3).
IL-6-572 and HMGB1 3814 did not show significant associations between genotypes or correlations with postictal serum levels in GEFS+ patients compared to controls (Table 3, Fig. 3C, D).
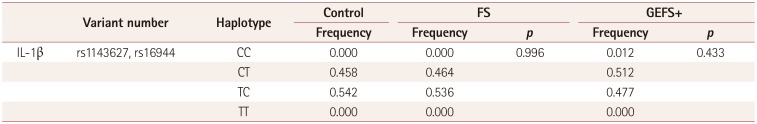
Haplotype frequency analysis of the block with IL-1β-31 and IL-1β-511
The haplotype frequencies in the block of IL-1β-31 and IL-1β-511 did not differ between FS and GEFS+ patients compared to controls (Table 4). CC and TT haplotypes at IL-1β-31 and IL-1β-511 were extremely rare (0–1.2%) and almost all patients had CT or TC haplotypes in the block of IL-1β-31 and IL-1β-511.
DISCUSSION
This is the first study showing proinflammatory cytokine IL-1β levels after an acute FS attack to be associated with genotypes of IL-1β-31 and IL-1β-511 promotor variants in children with FS. We found near-complete LD between the IL-1β-31 and IL-1β-511 variants, and the CT genotypes of both IL-1β-31 and IL-1β-511 showed significant correlations with higher postictal serum IL-1β levels compared to the CC+TT genotypes in children with FS. For GEFS+, CT genotypes of both IL-1β-31 and IL-1β-511 did not show significant association with postictal serum IL-1β after an AFS attack, probably being due to the small numbers of patients included. These data may support postictal serum IL-1β levels varying depending on the genotypes of IL-1β promotor variants in individuals. Therefore, serum IL-1β levels can be a critical factor determining whether or not FS develops at the time of a febrile state, representing evidence of genetic risk factors for fever-related seizures in children. However, we did not find a significant association between IL-6-572 and HMGB1 3814, which are known to be important proinflammatory cytokines related to FS and epilepsy.
IL-1β reportedly provokes seizures and also can be produced after seizures in animal models of epilepsy.18 IL-1β provokes seizures via up-regulation of excitatory neurotransmission19 and down-regulation of inhibitory GABA-mediated neurotransmission.20 Moreover, the intraventricular injection of IL-1β can induce FS even at a lower body temperature in 2-week-old mice during a febrile state.21 Seizure activity in itself triggers the production of cytokines in astrocytes and microglia, which are innate immune cells in the brain, even without peripheral immune cells or blood-borne mediators. IL-1β also enhances the main inflammatory responses and triggers the expression of other proinflammatory cytokine genes including IL-2, IL-6, IL-12, and TNF-α.22 Therefore, postictal IL-1β levels can be both a cause of seizures and a result of seizures. Although postictal serum IL-1β levels may reflect a result of seizures, higher sustained IL-1β levels also can trigger seizures.
The production of cytokines seems to change according to variants of cytokine genes.23 The action of promoter variants on gene transcription may be complicated and involve interactions with many variants sites. Therefore, genetic variants that up-regulate cytokine production can increase the degree of inflammatory responses to infection or other stimuli, which may subsequently predispose subjects to develop fever-provoked seizures. In vitro stimulation of peripheral blood mononuclear cells with lipopolysaccharide, mimicking bacterial infection, reportedly demonstrates that IL-1β production is higher in FS children than in controls.24
Several meta-analyses have investigated the associations of IL-1β promotor variants with the risk of FS. A recent meta-analysis studies showed that IL-1β-511 genotypes are associated with FS in both Asians and Caucasians.11 However, the genotypes with the strongest associations differed between Asians and Caucasians, with CT+TT vs. CC for Caucasian populations (OR=1.434) and TT vs. CC+CT for Asians (OR=1.393). These genetic differences may result from ethnic diversity or a lack of matching data on postictal IL-1β levels. There have been no previous reports on actual postictal IL-1β levels in FS with specific genotypes of the IL-1β-511 variant. In the present study, CT vs. TT+CC at IL-1β-511 showed significant correlations with higher postictal serum IL-1β levels, and the CT genotypes were associated with FS 1.731-fold more than were the TT+CC genotypes. The postictal serum IL-1β levels decreased in the following order of IL-1β-511 genotypes: CT>TT>CC. This suggests that CT and TT genotypes represent more-susceptible genetic variants in FS than CC genotypes in Asian children, which is consistent with the findings of a previous meta-analysis.11 The IL-1β-31 variant involves a TATA-box polymorphism that markedly affects DNA-protein interactions in vitro.25 T allele of IL-1β-511 reportedly up-regulates promotor activity, consequently increasing IL-1β production both in vitro and in vivo.926 The present study found near-complete LD between IL-1β-31 and IL-1β-511 variants, similar to a study of adult gastric cancer.25 If the genotype of IL-1β-31 is CT, the genotype of IL-1β-511 is also automatically determined as CT. The present study found that CT genotypes of both IL-1β-31 and IL-1β-511 were significantly associated with higher postictal serum IL-1β levels in children with FS. This finding may explain why the serum IL-1β level was identical in patients with CT genotypes of IL-1β-31 and IL-1β-511 in this study, because almost 99% of the patients carried CT or TC haplotype of IL-1β-31 and IL-1β-511. This situation made it impossible to evaluate the independent contribution of these variants to increased serum levels of IL-1β. Therefore, our results suggest that IL-1 gene cluster variants in IL-1β-31 and IL-1β-511 are a host genetic factor for provoking FS in Korean children.
Heterozygote genetic variants in membrane receptors may express multiple actions that affect the phenotypes. There are studies showing associations of heterozygotes of IL-1β-3954 (rs1143634) with posttraumatic epilepsy.27 In our results, heterozygotes of IL-1β promotor variants also showed significant associations with fever-provoked seizures and higher postictal IL-1β levels in children with FS. Different isoforms of IL-1β of different genotypes may influence receptor binding, transcriptional activation, and inflammation signaling pathways.28 Diverse variants in a single gene may be related to multiple genes and factors in the pathway responsible for the development of seizures. Therefore, understanding the role of each genetic variant is needed to explain their interactions.
GEFS+ is a familial epilepsy syndrome associated with FS and the subsequent development of epilepsy.3 However, the genetic mechanisms underlying the sensitivity to fever-related seizures in GEFS+ are currently unknown. GEFS+ patients in the present study showed similar patterns of genotypes and postictal serum IL-1β levels found in FS. CT genotypes of both IL-1β-31 and IL-1β-511 showed relatively higher postictal serum IL-1β levels than CC and TT genotypes after an AFS attack, although statistically insignificant. The lack of statistical significance may have been due to the small numbers of patients enrolled or the presence of AFS rather than FS. However, even AFS in GEFS+ was correlated with higher postictal serum IL-1β in patients with CT genotypes compared to TT+CC genotypes, suggesting that a genetic susceptibility to inflammation underlies seizure development.
HMGB1 is involved in the development and recurrence of seizures, and HMGB1 antagonists suppress the precipitation and recurrence of seizures in animal models of epilepsy.2930 HMGB1 is also found at higher levels in epileptogenic brain lesions of epilepsy patients.29 Extracellular HMGB1 binds to IL-1β and lipopolysaccharide and enhances inflammatory activities.3132 Serum HMGB1 was found to be significantly increased in children with FS and correlated with the serum IL-1β level.6 HMGB1 genes are highly conserved, with only seven variants being found and with an identical occurrence among various racial groups. The HMGB1 3814 polymorphism at intron 1 is known to affect the binding site of v-Myb, which is a powerful enhancer of HMGB1 expression.33 However, the HMGB1 3814 vatiant was not associated with either FS or GEFS+ in the present study.
IL-6 is a key proinflammatory cytokine involved in systemic inflammatory processes and is related to many neuroinflammatory disorders. The IL-6-572 C allele could increase the transcriptional activity of IL-6 promoter, leading to the up-regulation of IL-6 in stress or infection.34 The IL-6-572 C allele reportedly acts as a biomarker of small-vessel diseases of the brain in elderly patients, suggesting the presence of chronic neuroinflammation.35 The IL-6-572 CC genotypes were associated with higher serum IL-6 levels compared to GG genotypes among cerebral palsy children, implicating an association with chronic prenatal neuroinflammation,34 but no differences were seen in Taiwanese FS children.13 The present study also found no significant association of IL-6-572 with either FS or GEFS+.
The main limitation of this study was the small number of patients for whom acute serum samples were available after an acute seizure attack, which was due to the difficulty of obtaining such samples from young children with FS or GEFS+. The number of acute postictal serum samples was small especially in GEFS+ compared to FS, which might have resulted in the absence of statistical significance in GEFS+. Parents of GEFS+ families have previous experience of AFS attacks, which makes them less likely to bring their children to the emergency department. Therefore, a multicenter large-scale study involving subjects of multiple races is needed to confirm the correlations between postictal serum cytokine levels and genetic variants in order to identify the genetic risk factors for FS and GEFS+.
This is the first study demonstrating that proinflammatory cytokine IL-1β levels after an acute FS attack are associated with genotypes of IL-1β-31 and IL-1β-511 promotor variants in children with FS. Near-complete LD exists between the IL-1β-31 and IL-1β-511 variants, and CT genotypes of both IL-1β-31 and IL-1β-511 were significantly associated with higher postictal serum IL-1β levels compared to CC+TT genotypes in children with FS. These new data may indicate that promotor variants in IL-1β genes (and not IL-6 or HMGB1 genes) are a potential genetic risk factor for fever-related seizures in children.
 XML Download
XML Download