

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
Increasing of antibiotic resistance situations has raised an awareness around the world. A group of bacteria which had been recognized as threats of this century was named Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter species [1]. Recently, WHO announced 12 bacteria that posed the greatest threat to human health. Among those, carbapenem-resistant Pseudomonas aeruginosa, Extended-spectrum β-lactamase (ESBL)-producing carbapenem-resistant Enterobacteriaceae, and carbapenem-resistant Acinetobacter baumannii were critical priority [2]. P. aeruginosa also belongs to this group because its resistance mechanisms such as efflux pumps, loss of porins, and production of beta-lactamase enzymes [3]. The overexpression of MBLs is also one of resistance mechanisms frequently found in carbapenem-resistant P. aeruginosa especially in severe infection.
To discriminate differences between each bacterial strains, multilocus sequence typing (MLST) is now recognized as a common tool using seven housekeeping genes [4]. This method was firstly introduced in 1998 and demonstrated major advantages in both macro- and micro-epidemiology with moderate to high discrimination power over many methods [4]. MLST method has been used in many pathogenic bacteria including P. aeruginosa which was introduced in 2004 [5]. Sequence type (ST) 235, ST111 and ST175 were considered as the majority of P. aeruginosa clinical isolates [6]. In Asia, there were some studies reported MLST of MBL-producing P. aeruginosa, mainly from China and Japan [78910]. However, some studies showed the difference in ST prevalence, such as ST244 in China [789]. ST357 also demonstrated high prevalence as the second most ST [1011]. Even though MBLs producing P. aeruginosa have been isolated in Thailand, only one study identified P. aeruginosa isolates of ST235 harboring blaVIM-2 [12]. Therefore, the objective of this study was to investigate the dissemination traits of P. aeruginosa harboring MBLs including novel types of MBLs, blaIMP-65, collected from hospitals across Thailand.
Materials and Methods
1. Bacterial culture collection
Carbapenems-resistant P. aeruginosa clinical isolates were collected from patients in eight hospitals across five regions of Thailand with human ethical approval from Mahidol University Institutional Review Board (Certificate No. MU-IRB 2011/025-0102). All hospitals are tertiary or university hospitals. A total of 153 P. aeruginosa clinical isolates were characterized as carbapenems resistance among multidrug resistance. Multidrug resistance (MDR) criteria in this study was defined as non-susceptible to at least 3 of 5 drug groups which used in Pseudomonas infection treatment including including anti-pseudomonal penicillin (piperacillin), cephalosporin (ceftazidime), carbapenems (imipenem and meropenem), fluoroquinolone (ciprofloxacin), and aminoglycoside (gentamicin). Carbapenems resistance (CR) was defined by being non-susceptible to at least one carbapenem [1314]. The susceptibility of P. aeruginosa clinical isolates was confirmed in the research laboratory by the disc diffusion method according to the Clinical and Laboratory Standards Institute (CLSI) guidelines.
2. Phenotypic screening for metallo beta-lactamase
Phenotypic screening for metallo-beta-lactamase (MBL) enzyme was performed by diffusion method divided into 2 major methods which were double-disk synergy test (DDST) and combine disk test (CD) using EDTA as metallo beta-lactamase inhibitor [15].
3. Genotypic detection of metallo beta-lactamase gene and detection of integronI gene
Metallo beta-lactamase genes were divided into 3 groups for IMP-type MBLs and 2 groups for VIM-type MBLs depended on genotypic relationship of metallo beta-lactamase genes [16]. NDM-type MBLs were also included in this study. Each set of primers were designed to detect MBL genes in each group (Table 1). blaIMP-1, blaVIM-2 and blaNDM-1 were used as controls. Whole-cell lysates obtained from clinical isolates were used as DNA templates for IMP-, VIM- and NDM-type MBLs detection by PCR method. All IMP-type PCR products were further identified for specific types of IMP-gene by PCR with IMP-G, IMP-H and IMP-13 primers. All PCR products were submitted for sequencing. Nucleotide sequences were compared with sequence databases using BLAST (http://blast.ncbi.nlm.nih.gov/).
Table 1
List of primers designed in this study

IMP-AF, IMP-AR, IMP-BF, IMP-BR, IMP-CF, IMP-CR, VIM-DF, VIM-DR, IMP-GF, IMP-GR, IMP-HF, IMP-HR, IMP-HR, IMP-13F, IMP-13R, IMP-NF, IMP-NR, NDM-F, NDM-R, mutL-F, mutL-R, nuoD-F, and nuoD-R are primer name assigned for each group of detected genes.
IMP, imipenemase; VIM, verona integron-encoded metallo-β-lactamase; NDM, New Delhi metallo-beta-lactamase.
All CR-MDR P. aeruginosa were submitted for integronI detection by PCR method. The specific primers, Int1-F and Int1-R, of integronI gene were used as previously described [17].
4. Amplification of new allele of IMP- metallo beta-lactamases
IMP-N primers were designed for a full gene amplification. blaIMP-14 was used as template because new allele of MBLs present high similarity sequences to blaIMP-14. The new allele of metallo-beta-lactamase sequence was translated to protein sequence by BioEdit program. A full sequence of new metallo-meta-lactamase allele was submitted to assign as a novel allele at NCBI database. DNA Sequences of novel allele were analyzed and compared with similar sequences which were blaIMP-14, blaIMP-14a, blaIMP-32 and blaIMP-48.
5. Phylogenetic analysis
Nucleotide sequences of the novel blaIMP-65 gene (KY315991) and other IMP type genes were aligned based on iterative refinement method using MUSCLE version 3.8.31 [18]. Phylogenetic tree of IMP type genes was reconstructed based on the randomized accelerated maximum likelihood. General time reversible model (GTR) with gamma distribution (G) and invariant sites (I) was a selected DNA substitution model for the phylogenetic analysis using RAxML version 8 [19]. The confidence values on phylogenetic tree were presented as percentage of bootstrap values (1,000 bootstrap replicates).
6. Multilocus sequence typing for metallo beta-lactamase producing P. aeruginosa
MBL harboring P. aeruginosa were selected as representatives when clinical isolates harbored the same MBL gene and demonstrated the same PFGE pattern [14]. A total of 14 clinical isolates P. aeruginosa was chosen and characterized for sequence typing (ST) by MLST method. Seven meropenem non-susceptible among MDR P. aeruginosa clinical isolates were chosen for comparison.
P. aeruginosa clinical isolates were characterized for molecular typing. MLST was performed as described previously with some modifications [5]. Briefly, seven housekeeping genes (ascA, aroE, guaA, mutL, nuoD, ppsA, and trpE), were amplified and sequenced. The nucleotide sequences were determined and compared with MLST database to assign the allele of these genes and STs (http://pubmlst.org/paeruginosa/). Primers for mutL and nuoD genes were modified in this study as showed in Table 1.
Results
1. Detection of integronI and metallo beta-lactamase genes
All CR-MDR P. aeruginosa were identified for phenotypic resistance pattern by diffusion method and genotypically detected for intI gene by PCR. One hundred and thirty six (88.9%) clinical isolates of P. aeruginosa were positive for int1 gene. Phenotypic screenings with EDTA were used to detect the presence of metallo beta-lactamases. One hundred and four clinical isolates (68.0%) of CR-MDR P. aeruginosa were positive by CDT and/or DDST. These strains were further subjected for metallo beta-lactamase genes detection by PCR with specific primers. Total of 31 (20.3%) P. aeruginosa clinical isolates were found to present metallo beta-lactamase genes which could be divided into VIM-type for 14 strains and IMP-type for 17 strains. For VIM-type, all 14 strains were detected by VIM-D primers and PCR products were sequenced and identified as blaVIM-2. However, NDM-type MBL could not be detected in this study.
For IMP-type MBLs, all 17 strains were detected by IMP-C primers. Those strains were amplified with IMP-G, IMP-H and IMP-13 primers to identify specific types of IMP-MBLs. PCR products were sequenced and found 8 blaIMP-1, 6 blaIMP-14 and only one blaIMP-13 harboring strains. However, two strains were carried unidentified MBLs.
2. Novel allele of IMP-type MBLs
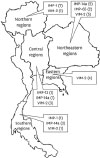
Those two strains carried unidentified IMP-type MBLs were further investigated. IMP-14 primers were designed based on blaIMP-14 sequence because unidentified MBLs could present high similarity sequences to blaIMP-14. The novel gene was mutated at position 279 and 301 from blaIMP-14 gene. Therefore, translated protein was changed from threonine (T) to alanine (A) at position 101 (Fig. 1). The novel gene was deposited and assigned as blaIMP-65 by the NCBI (accession number KY315991). Dissemination of MBL genes detected in P. aeruginosa across Thailand was showed in Fig. 2. Translated protein sequences of IMP14, IMP-14a, IMP-32, IMP-48 and IMP-65 were compared as showed in Fig. 1. These sequences presented high similarity with one or two amino acids change.

3. Phylogenetic Analysis
The phylogenetic tree represented the relationship among the novel IMP-type gene (blaIMP-65) and other IMP-type genes (Fig. 3). There were three groups of the IMP-type genes (IMP-type group I, II and III) presented on the tree. The novel IMP-type gene was affiliated with the IMP-type group I. blaIMP-32, blaIMP-48, blaIMP-14a, blaIMP-14 and blaIMP-54 were the closest relative genes of blaIMP-65. This IMP-type subgroup had been supported by 100% of bootstrap (Fig. 3). It was clearly demonstrated that Thailand was the origin of blaIMP-14, blaIMP-14a, blaIMP-48 and blaIMP-65 isolates (Supplementary Table 1).
Figure 3
Evolutionary relationships of IMP-type genes. The phylogenetic tree reconstructed using the randomized accelerated maximum likelihood method with the GTR+I+G model of DNA substitution. The values associated with nodes correspond to the bootstrapping support in %. Bootstrap value lower than 50% was not represented on the clades.
IMP, imipenemase; GTR, general time reversible model; I, invariant sites; G, gamma distribution; DNA, deoxyribonucleic acid.

4. MLST profile analysis
Based on MLST analysis, a total of 11 clinical isolates of MDR P. aeruginosa in Thailand were identified as ST235 (Table 2). Among those, six clinical isolates were MBLs-producing P. aeruginosa which harbored blaVIM-2 for 5 clinical isolates and only one isolate harbored blaIMP-14a. All blaIMP-1 producing isolates were identified as ST964. Moreover, both blaIMP-65 harbored P. aeruginosa were characterized as ST111. Others MBLs harboring P. aeruginosa were identified as ST292, ST621 and ST1584 which were found only one clinical isolate each. Interestingly, most blaVIM-2 carrying P. aeruginosa were identified as ST235 but one isolate was identified as ST1584. Among non-harbored MBL gene CR-MDR clinical isolates, ST235 was also identified as a dominant type. ST273 and ST1816 were also identified in this non-harbored MBLs group. It was noteworthy that ST235 was found as dominant ST (52.4%) in Thailand followed by ST964 (14.3%) and ST111 (9.5%). ST292, ST273, ST621, ST1584 and ST1816 were found only in one clinical isolate each.
Table 2
Multi-locus sequence typing allelic profiles and sequence types of 21 Pseudomonas aeruginosa clinical isolates collected across Thailand

S, susceptible; I, intermediate; R, resistant; IPM, imipenem; MEM, meropenem; PIP, piperacillin; CAZ, ceftazidime; CIP, ciprofloxacin; GEN, gentamicin; ST, sequence typing.
A-G, indicated similar PFGE fingerprinting; ND, no ST determined; N, northern region; NE, northeastern region; C, central region; E, eastern region; S, southern region.
Discussion
In our study, clinical isolates were phenotypic screened by paper disc diffusion, DDST and CDT. Many clinical isolates were interpreted as positive. However, only 20% were detected MBL genes. These might be affected by the obscure definition of DDST method which was the appearance of either an enhanced or a phantom zone between the antimicrobial agent and the inhibitor disk was considered a positive result and indicative of MBL production. Moreover, IntegronI was also found in most tested clinical isolates and always related with many antibiotic resistance mechanisms. These might indicate that those CR-MDR clinical isolates could carry many antibiotic resistance mechanisms such as, loss of OprD, multiple efflux pumps, or antibiotic inactivating enzymes and could be affected the phenotypic screening method. However, all positive phenotypic screened were further identified for specific type of MBLs.
Most prevalence MBLs belong to IMP- and VIM- type in P. aeruginosa. However, one of the most recent discovered MBLs, the NDM-type was also reported worldwide [20]. In our studies, we discovered blaIMP-1, blaIMP-13, blaIMP-14a, blaVIM-2 and a novel MBLs, blaIMP-65. Previous report of VIM-type MBLs in Thailand were blaVIM-2, blaVIM-5 and blaVIM-45 [21]. In this study, we found only blaVIM-2 but these clinical isolates were collected from tertiary or university hospitals located in different parts of Thailand. This might indicate that blaVIM-2 MBLs could be dominated VIM-type MBLs in Thailand.
Some previous reports showed a variety of IMP-type in Thailand including blaIMP-1, blaIMP-13, blaIMP-14, blaIMP-14a, blaIMP-32 and blaIMP-48 [2122232425]. In this study, many IMP-type MBLs sush as blaIMP-1, blaIMP-13, blaIMP-14a and blaIMP-65 were identified. The results also indicated that blaIMP-1 and blaIMP-14a might be dominant types of IMP-MBLs in Thailand. Novel IMP-type MBLs, IMP-65, was demonstrated an amino acid changed from IMP-14 at the position 101 from alanine to threonine. There were also some IMP-type, blaIMP-32 and blaIMP-48, which presented similar sequences with blaIMP-65 and blaIMP-14. Moreover, blaIMP-14, blaIMP-14a, blaIMP-32, blaIMP-48 and blaIMP-65 were never reported in any clinical isolate from other countries than Thailand. These might indicate that blaIMP-14 and variants were spreading locally and also dominant in Thailand.
Acquired MBL genes were always found within class I integrons which usually carried other additional resistance genes and also known as a gene cassette. IntegronI is an important mechanism that bacteria use to transfer resistance mechanisms. Most class I integrons are always carried within transposons which help the bacteria to transfer antibiotic resistance gene from one to another [26]. In this study, we also found that most MDR P. aeruginosa carried integronI gene. Moreover, all MBLs harbored P. aeruginosa except one strain (96.6%) also carried integronI gene which was similar to many previous studies [2728].
The primers used in this study were designed to target different alleles of IMP-type MBLs. All IMP-type MBLs in Thailand were detected by IMP-C primers designed to detect all previously report IMP-type MBLs in Thailand but it was unable to distinguished specific alleles of IMP-type MBLs in Thailand. From these results, this set of primers may be used as molecular confirmation test to detect dominant type of IMP-type MBLs type for epidemiology control in Thailand.
ST235 was identified as a dominant ST in Thailand. Moreover, we also found that ST964 was also presented as a significant ST type among MBL-harbored P. aeruginosa. However, this study was emphasized that ST235 was the dominant ST in Thailand when compared the results with previous studies [1229]. Kim et al. studied dissemination of carbapenemase P. aeruginosa but only a few clinical isolates from Thailand were included and found only ST235 [12]. Another ST from Thailand is ST260 which was detected in P. aeruginosa harbored blaIMP-14 [29].
ST235, ST111 and ST175 are high-risk clones which now concerned as majority of P. aeruginosa clinical isolates [6]. However, in Asian countries, only a few ST111 have been reported [8]. In this study, ST111 was identified but ST175 was undiscovered. ST111 were also carried the novel IMP-type MBL, blaIMP-65. These finding was quite interesting since ST111 was occasionally found in Asia and blaIMP-65 or similar IMP-types have never been reported in other places. Unfortunately, the information about those strains cannot be retrieved other than the location of the collection.
ST273, ST292, ST621, ST964, ST1584, and ST1816 have also never been reported in Thailand before. ST964, the second most ST in this study, was reported in Singapore and UK [3031]. Those stains of ST964 in Singapore were also carried blaIMP-1 as found in our study. ST292 was another ST which were reported in Asian countries, China and Korea [3233]. However, there was no report on ST621, ST1584, and ST1816 in Asia. It was interesting that we found ST621 which was always carrying blaIMP-13 as same as in many countries [34]. From these results, it might indicate the different dissemination trait from European countries which identified a lot of ST111 and ST175. However, the larger groups of samples may be required to gain more data.
It was noteworthy that antibiotic susceptibility in this study was determined by disc diffusion method according to the CLSI guideline. Therefore, some alternative drugs such as, colistin, were not examined due to unavailable breakpoints. In addition, only strains shared similar PFGE fingerprinting were selected as representative strains
In conclusion, this was the first report on blaVIM-2 and blaIMP-1 types which were highly presented MBL genes in Thailand. Moreover, the novel IMP type, blaIMP-65 was also identified and dominated locally in MBLs, blaIMP-14 and variants. ST235 was dominant among MBL-producing P. aeruginosa as well as no MBL-producing P. aeruginosa while each of ST964 and ST111 represented the clones of P. aeruginosa producing IMP-1 and IMP-65, respectively. Moreover, it is also the first report on many P. aeruginosa STs in Thailand: ST273, ST292, ST621, ST1584, and ST1816 which emphasized the dissemination trait difference of MBLs harboring P. aeruginosa in Thailand.
 XML Download
XML Download