

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Gastric cancer (GC) is becoming more common all around the world. In fact, it is believed to be the third most common of all cancers among males, as well as the fifth most common among females.1 Lower than 20% of GC patients can survive for more than 5 years after diagnosis. The incidence of GC has reduced in developed countries, whereas it is increasing in developing countries.2 Helicobacter pylori (HP) is a gram-negative bacterium that plays an important role in the pathogenesis of GC.3 About 60% of GC patients in developed countries and 75% in developing countries have chronic infection of HP.4 Chronic gastritis caused by HP infection leads to the release of pathogenic factors like urease, vacuolating toxin A, CagA protein, inflammatory mediator, and reactive oxygen metabolites, causing aberrant hyperplasia and apoptosis of gastric mucosal epithelial cells, eventually leading to GC.56 Despite recent advances in treatments, including surgery, chemotherapy, and radiation therapy, GC still remains a difficult cancer to cure.7 Therefore, to cure GC, certain molecular mechanisms underlying GC cells proliferation, invasion, and migration, especially in HP-infected GC must be elucidated.
MicroRNAs (miRNAs) are endogenous noncoding RNAs with 17–25 nucleotides, which play considerable roles in gene regulation of pathogenesis.8 Recent studies have confirmed that miR-1915 is related to several types of cancer, such as hepatocellular cancer,9 lung cancer,10 breast cancer,11 and colorectal cancer.12 Importantly, miR-1915 was down-regulated in GC cells,13 which might be major miRNA targets deserving further investigation. Nevertheless, the molecular mechanisms of miR-1915 trigger HP-infected GC remain elusive.
Receptor for advanced glycation end product (RAGE) is a member of the immunoglobulin superfamily, which consists of more than 400 amino acids with a molecular weight of 35 kD. RAGE has been shown to be linked with poor prognosis in GC patients,14 and our early study has also proven that knockdown of RAGE inhibited growth and invasion of GC cells.15 In addition, RAGE is over-expressed in GC tissues, especially in HP-infected GC cells.16 These data indicated that RAGE may be strongly linked to the pathogenesis of HP-infected GC, although the mechanisms behind disease progression remain unknown. Notably, we found that miR-1915 and RAGE sequences have binding sites by using bioinformatics software (microRNA.org), which demonstrated that RAGE may be a downstream target molecule of miR-1915.
In this context, we tried to clarify whether miR-1915 is causally involved in GC with HP infection, by performing several systematic and bioinformatic approaches and studying GC cell lines and GC tissues from human stomach biopsy specimens. Furthermore, our work aims to define the molecular mechanisms between miR-1915 and RAGE in regulation of cell proliferation, invasion, and migration of HP-infected GC cells.
MATERIALS AND METHODS
Human tissue specimens
This study on human beings has been approved by the Institutional Review Board (IRB) of The First Affiliated Hospital of Xinjiang Medical University. For the role of materials for research purposes, written informed consent was received from each patient. Human tissue specimens were accumulated and classified as described.17 Twenty pairs of H. pylori-positive gastritis [HP (+) Gastritis] and negative gastritis [HP (−) Gastritis] tissues were obtained from patients who underwent gastroscrope in The First Affiliated Hospital of Xinjiang Medical University. Thirty GC tissues were obtained randomly from patients who underwent gastrectomy in the same hospital. All GC tissues were identified by rapid urease test, and positive specimens were assigned to H. pylori-positive GC [HP (+) GC] group, while negative ones were assigned to H. pylori-negative GC [HP (−) GC] group. There were 16 specimens in HP (+) GC group and 14 specimens in HP (−) GC group. All tissue specimens were formaldehyde-fixed paraffin-embedded.
Cell lines, H. pylori strain and HP infection
Human normal gastric epithelial cell line (GES-1) was bought from Shanghai Institute of Cell Biology (Shanghai, China). Human GC cell lines (SGC-7901 and MKN45) and wild-type H. pylori strain 26695 were bought from American Type Culture Collection (ATCC, Manassas, VA, USA). Infection procedure was performed as described.18 Cell lines SGC-7901 and MKN45 were propagated in DMEM (Gibco, Invitrogen, Waltham, MA, USA) with 10% FBS in a humidified incubator (5% CO2 at 37℃). After cells grew to be approximately 80% confluent, they were co-cultured with HP at multiplicity of infection (M.O.I) of 100:1. After 12 h of infection, total RNA was extracted. All cell lines were maintained in RPMI 1640 that was supplemented in a humidified atmosphere of 95% air and 5% CO2 with 10% fetal calf serum at 37℃, which were identified by authentication.
Quantitative real-time PCR
Total RNAs were extracted from gastric tissues or cell lines using TRIZOL reagent (Invitrogen, Carlsbad, CA, USA). Reverse Transcription Kit (Takara, Dalian, China) was used to reverse transcribe from RNA to cDNA. The expressions of miRNAs and mRNAs were analyzed by quantitative real-time PCR (qRT-PCR) using Power SYBR Green (Takara) with U6 or GAPDH as endogenous controls, respectively. For detection of miR-1915 expression, reverse transcription was performed as Applied Biosystems TaqMan MicroRNA Assay (Foster City, CA, USA) protocol. The RNA sequences used in this study were as follows: miR-1915: (Forward) 5′-CCCAAGCTTGGAAATCCGACCACTA-3′, (Reverse) 5′-CATGCCATGGCAGGATAGCAGCAC-3′; RAGE: (Forward) 5′-GTGTCCTTCCCAACGGCTC-3′, (Reverse) 5′-ATTGCCTGGCACCGGAAAA-3′; β-actin: (Forward) 5′-TGATCCACATCTGCTGGAAGGT-3′, (Reverse) 5′-GACAGGATGCAGAAGGAGATTACT-3′. Relative expression levels of all genes were calculated as 2−ΔΔCt.
Western blot analysis and antibodies
Total proteins were extracted from specimens or cell lines by 10% SDS-poly acrylamide gel electrophoresis, and transferred to poly-vinylidene difluoride membranes (Millipore, Burlington, MA, USA). After blocking with 5% fat-free milk in 1×TBST for 1 h, total proteins were immunoblotted with specific antibodies at 4℃ overnight. After three times of washing, the proteins were incubated with horseradish peroxidase-conjugated secondary antibodies for 1 h at room temperature. After another three times of washing, protein bands were detected by SmartChemi (Beijing Sage Creation Science Co, Beijing, China). Antibody β-actin was used as internal reference.
Cell transfection
The 293T cells were bought from Shanghai Institute of Cell Biology. The cells (2×104 cells/well) were cultured in 24-well plates overnight, and transiently transfected using transfection reagent lipofectamine 2000 (Invitrogen). MiR-1915 mimic and negative control mimics (pre-NC), miR-1915 inhibitor, and negative control inhibitor (NC) were purchased from RiboBio Co., Ltd. (Guangzhou, China).
Cell counting kit-8 (CCK-8) assay
The 7901 or MKN45 cells (2×103 cells/well) were cultured in 96-well plates in 100-µL culture medium overnight. CCK-8 reagent (10 µL/well) was added, and they were incubated for another 4 h with 5% CO2 at 37℃. We observed and recorded the procedures of cell growth at 0, 24, 48, and 72 h. Cell viability was determined at 450 NM, and expressed as a percentage of the control using a microplate reader (Bio-Rad, Hercules, CA, USA).
Migration and invasion assay
Capability of cell migration was examined by transwell migration assays. Transwell migration chambers were performed. Cells were seeded to 1×105 cells in serum-free media in the upper chamber with noncoated membrane (8 µm in pore size; Millipore, Schaffhausen, Switzerland). Lower chamber contained media with 20% FBS as a chemoattractant. Cells in upper chamber were discarded by using cotton wool after 24 h, and migration cells in lower chamber were counted by microscope. For invasion assay, the experiments were the same as transwell migration assays, except that cells grew to 2×105 cells in upper chamber, which was coated with Matrigel (Sigma, St. Louis, MO, USA).
Luciferase reporter assay
We used TargetScan Human Release 6.2 (http://www.targetscan.org) to determine predicted target genes as well as their binding sites. The 3′-UTR sequence of RAGE was a candidate target gene of miR-1915, and was inserted into the pmirGLO Dual-Luciferase miRNA Target Expression Vector. The 293T cells were plated in 24-well plates and co-transfected with the vector carrying wild-type (WT) or mutated (Mut) RAGE 3′-UTR and miR-1915 mimic or the negative control (NC). After 48 h of transfection, luciferase activity was detected by luciferase reporter assay system (Promega, Madison, WI, USA).
Statistical analysis
SPSS version 18.0 (SPSS Inc, Chicago, IL, USA) was used for statistical analysis, with either a Student's t-test or analysis of variance. All experiments were performed in triplicate, and data were presented as mean±standard deviation. p-values less than 0.05 were considered statistically significant.
RESULTS
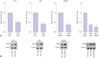
Lowly-expressed miR-1915 and over-expressed RAGE are detected in HP (+) GC tissues in vivo
MiRNAs and RAGE have been shown to be causally involved in gastric malignancy.1315 We assumed that cell proliferation, invasion, and migration of HP (+) GC cells were promoted by molecular regulation between them. We first examined miRNA expression levels in HP-infected and uninfected GC tissues. We gathered 20 pairs of gastritis specimens as control, and 30 GC specimens with 16 HP (+) and 14 HP (−) specimens. Several miRNAs, such as miR-20b,19 miR-342,20 miR-577,21 miR-767-5p,22 and miR-1915,13 were believed to participate in gastric tumorigenesis, and their expressions in GC tissues were detected by qRT-PCR. Only the expression of miR-1915 was significantly reduced in HP (+) or HP (−) GC groups compared to the control groups, while other miRNAs expressions were increased (Supplementary Fig. 1, only online). Among them, miR-1915 was found to be an intriguing candidate, as it played an important role in GC (significant lowly-expressed in GC tissues).
To determine the expression of miR-1915 and RAGE during the development of HP-associated GC in vivo, we further studied gastric tissues from patients with or without HP infection. The qRT-PCR results showed that the expression of miR-1915 was down-regulated in HP (+) Gastritis and GC groups compared to HP (−) groups. Additionally, miR-1915 expression remained under-expressed in HP (+) GC group compared to HP (+) Gastritis group (Fig. 1A). For the expression of RAGE, western blot analysis showed that RAGE expression was increased in HP (+) Gastritis and GC groups compared to HP (−) groups, and remained over-expressed in HP (+) GC group compared to HP (+) Gastritis group (Fig. 1B).
Lowly-expressed miR-1915 and over-expressed RAGE are detected in HP (+) GC cells in vitro
To examine the in vitro expression of miR-1915 and RAGE during the development of HP-associated GC, GES-1, SGC-7901, and MKN45 cell lines were obtained and infected with H. pylori at M.O.I of 100:1. Total RNA was extracted 12 h after H. pylori infection. Before the infection, expression of miR-1915 was reduced and expression of RAGE was enhanced in HP (−) 7901 and MKN45 cells compared to HP (−) GES-1 cells (Supplementary Fig. 2, only online). After H. pylori infection, qRT-PCR results showed that miR-1915 expression was reduced in HP (+) GES-1, 7901, and MKN45 cells compared to their HP (−) controls. Moreover, expression of miR-1915 remained under- expressed in HP (+) 7901 and MKN45 cells compared to HP (+) GES-1 cells (Fig. 2A). In contrast, western blotting results showed that RAGE expression was increased in HP (+) GES-1, 7901, and MKN45 cells compared to their HP (−) controls, and remained over-expressed in HP (+) 7901 and MKN45 cells compared to HP (+) GES-1 cells (Fig. 2B). Together, these data demonstrate that miR-1915 is lowly-expressed, but RAGE is over-expressed in HP (+) GC cells in vivo and in vitro.
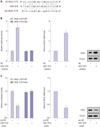
Over-expressed miR-1915 inhibits proliferation, invasion, and migration of HP (+) GC cells
Given the lowly-expressed miR-1915 in HP (+) GC cells, we sought to elucidate the causative effect in cell proliferation, invasion, and migration. The HP (+) 7901 and MKN45 cells were transfected by miR-1915 mimic, thus the expression of miR-1915 was increased (Fig. 3A). Then GC cell proliferating capability was examined by CCK8 assay. We observed the procedures of cellular growth in 0, 24, 48, and 72 h, and found that GC cell proliferate capability was attenuated in miR-1915 mimic group compared to pre-NC group (Fig. 3B). GC cell invasion and migration capability were also examined in HP (+) 7901 and MKN45 cells by transwell assays, and the results showed a decline in miR-1915 mimic group (Fig. 3C). Taken together, these data demonstrate that up-regulation of miR-1915 inhibits cell proliferation, invasion, and migration of HP (+) GC cells.
There is a target relationship between miR-1915 and RAGE
To achieve the molecular regulation pattern between miR-1915 and RAGE, we introduced miR-1915 inhibitor into 293T cells. Software predicted that miR-1915 could bind to 3′UTR region of RAGE (Fig. 4A). Luciferase reporter assay showed that the relative luciferase activity of RAGE 3′UTR (WT) was significantly increased. Additionally, RAGE mRNA level and RAGE protein expression were also increased, as detected by qRT-PCR and western blot analysis (Fig. 4B). We also introduced miR-1915 mimic into 293T cells, and thus the relative luciferase activity of RAGE 3′UTR (WT) was significantly reduced. The qRT-PCR and western blot analysis results revealed that RAGE mRNA level and RAGE protein expression were both reduced (Fig. 4C). Therfore, we could prove that there is a target relationship between miR-1915 and RAGE.
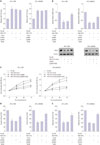
Molecular mechanism of HP (+) GC cell proliferation, invasion, and migration via miR-1915
To confirm the specific molecular mechanisms mediated by miR-1915, HP (+) 7901 and MKN45 cells were transfected with miR-1915 mimic and treated with pcDNA-RAGE, and corresponding control groups were established. The qRT-PCR results showed that miR-1915 level was significantly increased after transfected by miR-1915 mimic compared to pre-NC group, while it remained unchanged after being treated with pcDNA-RAGE compared to pcDNA group (Fig. 5A). On the other hand, RAGE mRNA level and RAGE protein expression were under-expressed after being transfected by miR-1915 mimic, and they remained over-expressed after being treated with pcDNA-RAGE in HP (+) 7901 and MKN45 GC cells (Fig. 5B).
Speaking of cell proliferation, invasion, and migration, HP (+) 7901 and MKN45 cells were transfected as previously described, and examined by CCK-8 and transwell assays. The results revealed that cellular capabilities of proliferation, invasion, and migration were all inhibited after transfected by miR-1915 mimic, but were promoted after being treated with pcDNA-RAGE (Fig. 5C, D, and E). Based on these results, we conclude that over-expressed miR-1915 modulation of under-expressed of RAGE may be the mechanism which attenuates the proliferation, invasion, and migration of HP (+) GC cells.
DISCUSSION
GC is a fatal disease, which is also known to be the second most common cause of malignant neoplastic disease-related deaths worldwide,23 and it is still difficult to cure this condition. The mechanisms by which HP-associated gastritis develops into GC is multifactorial. HP infection induces expression of proinflammatory cytokines, such as interleukin (IL)-8 and tumor necrosis factor (TNF)-α, by transactivation of SRE and AP-1 in GC.6 HP infection also activates the cyclin D1 gene through mitogen-activated protein kinase (MAPK) pathway and overexpression of cyclin D1 accelerates cell aberrant proliferation.5 In addition, activation of telomerase, ras, c-met, c-myc, and c-erbB-2 genes, as well as the inactivation of p53 gene are involved in HP-associated gastric malignant tumorigenesis.9 To this end, we can conclude that HP infection significantly increases the incidence of GC, and clarify that the mechanisms between them are particularly important for treatment of GC.
Unlike tumor-suppressor-miRNAs, oncomiRNAs are usually over-expressed in GC, and they promote the proliferation of GC cells, which is a causally step in cancer development. MiR-199a/b-3p,24 miR-21,25 miR-130b,26 and miR23a27 were confirmed to be oncomiRNAs promoting GC cell proliferation. However, over-expression of tumor-suppressor-miRNAs leads to the slowdown of cancer cell growth.28 Jin, et al.29 found that miR-582-5p suppressed GC cell proliferation via targeting AKT3. The expressions of miR-181c,30 miR-212,31 and miR-51232 were silenced with DNA hypermethylation in GC, and their restored expressions could induce reduced GC cell growth through inhibition of oncogenes expression. Aside from cell proliferation, cell invasion and migration are both important indicators for assessing the malignancy of cancer cells. Ectopic expression of miR-101 has been proven to inhibit cell migration and invasion of GC cells via mediating MCL-1, FOS, EZH2, and COX-2 genes.33 Down-regulation of miR-335 was causally related to metastasis of GC lymph-node, as well as invasion of lymphatic vessels.34 Although an accumulation of studies on onco-related miRNAs were performed in recent years, most of them focused on GC cells rather than HP-infected GC cells. Our current study confirmed that miR-1915 exerted tumor-suppressive effects on HP (+) GC cell proliferation, invasion, and migration, which provided an innovative and candidate target for treatment of HP-infected GC. Before H. pylori infection, expression of miR-1915 was reduced while expression of RAGE was enhanced in HP (−) 7901 and MKN45 cells compared to HP (−) GES-1 cells (Supplementary Fig. 2, only online). Combined with the down-regulation of miR-1915 in HP (−) GC tissues, these data indicated that miR-1915 was down-regulated in both HP (−) GC tissues and cells; therefore, their underlying mechanisms deserve further investigation in our future research.
Recent studies have confirmed that miR-1915 is related to carcinogenesis. For instance, miR-1915-3p has been identified as oxidative stress-responsive miRNA regulated by p53-dependent pathway in antiapoptosis process in hepatocellular cancer.9 Such antiapoptosis function of miR-1915-3p has also been identified in lung cancer by targeting GTP-binding protein 2 (DRG2)/pre-B cell leukemia homeobox 2 (PBX2).10 Furthermore, miR-1915-3p enhances breast cancer cell proliferative and migrational abilities by targeting gene DUSP3 and activating ERK1/2.11 On the contrary, our work revealed the negative effect of miR-1915 in cell proliferation, invasion, and migration of HP-infected GC cells by targeting RAGE. We assumed that this may be associated with the infection of HP and immunologic function of RAGE. Taken together, these data have revealed that miR-1915 functions as either oncogenes or tumor suppressor genes depending on the roles of its target gene, which emphasizes the importance of miR-1915 in different cancer treatments.
The above mentioned mechanism needs an important molecule participation—RAGE. It is a pattern recognition receptor which also plays an important role in the occurrence of GC via binding multiple ligands derived from a damaged cell environment.35 RAGE has previously been shown to be related to invasion and metastasis in GC,36 which indicates a research base for the current study of RAGE in GC. To this end, our former study focused on RAGE in GC, and proved that knockdown of RAGE could inhibit growth and invasion of GC cells.15 Inspired by our former research, we assumed that targeted inhibition of RAGE may act as novel targets to improve current cancer therapies. In this study, we detected RAGE expression in HP-infected GC tissues and cells by using several systematic and bioinformatic approaches, and first found that RAGE expression could be reduced by up-regulating miR-1915 by which cellular proliferation, invasion, and migration were inhibited in HP-infected GC cells. Similarly, the role of RAGE in other cancer types, by promoting cell proliferation and tumor metastases, has been reported. For instance, inhibition of RAGE decreased cell proliferation of breast cancer cells,37 induced cell apoptosis, and inhibited growth of prostate cancer.38 RAGE dysfunction inhibited colorectal cancer angiogenesis and its progression as well,39 and prolonged the survival rate of patients who were diagnosed with pancreatic cancer.40 In addition to the relationship between miR-1915 and various types of cancers, miR-1915/RAGE pathway in other types of malignant cells deserves further investigation.
In conclusion, we confirmed that miR-1915 exerted tumorsuppressive effects on HP-infected GC cellular proliferation, invasion, and migration via targeting RAGE, which may be a novel prognostic biomarker and a potential therapeutic target for HP-infected GC patients.


 XML Download
XML Download