

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Dendritic cells (DCs) are the most potent antigen presenting cells, which are crucial for the induction of T cell responses.12 DCs can acquire and process antigens in the periphery, and migrate to secondary lymphoid tissues where they prime primary T cell responses. While DCs play a key role in the initiation of primary immune responses, they also play a crucial role in the development and maintenance of immune tolerance.345
The functional difference between immunogenic and tolerogenic DCs depends on maturation state and maturation environment. Immature tissue-resident DCs sense invading antigens via pattern-recognition receptors such as toll-like receptors, take up the antigens via phagocytosis or endocytosis and degrade them into small peptides to present the antigenic peptides in association with major histocompatibility complex (MHC) class II molecules to CD4 T cells.1 These antigen entrapping and processing processes trigger the maturation of immature DCs to mature DCs, which express considerably higher levels of co-stimulatory molecules such as CD80 and CD86, and MHC class II molecules, and secrete considerably higher amounts of proinflammatory cytokines, such as interleukin (IL)-1, IL-6, IL-12, and tumor necrosis factor (TNF)-α, compared to immature DCs.6
Tissue-resident steady-state DCs are immature DCs, which express low levels of co-stimulatory molecules and moderate levels of MHC class II molecules, and are poorly immunogenic unless activated. In fact, steady-state immature DCs, which display peptides originating from self-proteins in association with MHC molecules on the cell surface, are tolerogenic DCs (tolDCs) that maintain self-tolerance against self-antigens.7 A number of attempts have been made to use tolerogenic immature DCs to induce immune tolerance. Dhodapkar and Steinman generated immature DCs using IL-4 and granulocyte macrophage-colony stimulating factor (GM-CSF), pulsed them with antigen, and then injected them into humans. They showed that injection of antigen-pulsed immature DCs led to antigen-specific inhibition of effector T cell function by inducing regulatory T cells (Tregs).89 However, using tolerogenic immature DCs to induce immune tolerance raises concerns regarding the functional stability of the immature state, because immature DCs could be converted into immunogenic mature DCs when encountering a ‘danger signal’ such as proinflammatory cytokines and microbial products. Thus, one of the major challenges facing tolDC-based immunotherapy is optimizing the protocol for obtaining functionally stable tolDCs.
TolDCs with durable immaturity and immune regulatory properties have been generated ex vivo using various pharmacological agents such as rapamycin, dexamethasone, and vitamin D.10 Immunosuppressive cytokines such as IL-10 and transforming growth factor (TGF)-β have also been used to induce tolDCs.11 In general, tolDCs are characterized by reduced expression of co-stimulatory molecules and IL-12, decreased ability to induce T cell proliferation, increased IL-10 secretion, and increased Treg induction.1011 The mechanisms underlying tolDC activity include the induction of Tregs, increasing the expression of programmed death-ligand 1 (PD-L1) and inducible costimulator ligand (ICOSL), and the production of immunosuppressive factors such as IL-10 and TGF-β.711121314
Antigen-pulsed tolDCs are promising tools for generating antigen-specific immune tolerance. They can be infused directly for the induction of antigen-specific immune tolerance in vivo, or can be used to generate antigen-specific Tregs in vitro for Treg-based adaptive cell therapy. In this review, we describe the methods used to generate tolDCs ex vivo and the phenotypic and functional characteristics of the induced tolDCs. In addition, we discuss the therapeutic potential of tolDCs for treating immune disorders based on completed or currently on-going clinical trials with tolDCs.
EX VIVO GENERATION OF tolDCs
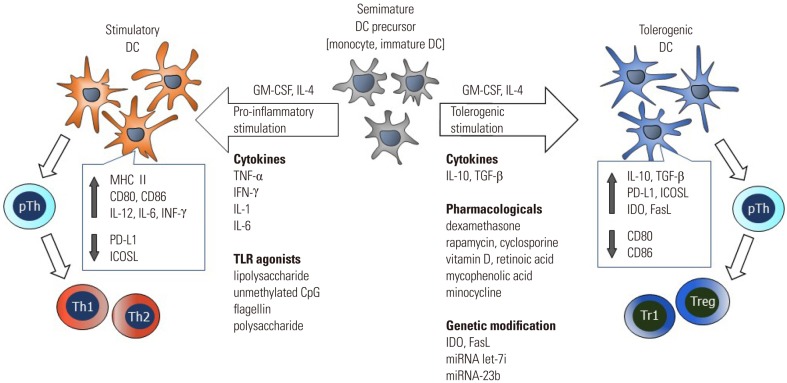
Human tolDCs are mostly produced from peripheral blood monocytes by culturing in the presence of GM-CSF and IL-4 together with an agent(s) known to confer tolerogenic properties. In murine systems, immature DCs are first generated by culturing bone marrow cells in the presence of GM-CSF and IL-4, and then induced to tolDCs by additional culturing in the presence of an agent(s) known to confer tolerogenic properties.15 Several pharmacological and biological agents have been used to generate tolDCs ex vivo from hematopoietic precursors or peripheral blood monocytes. The major methods used to generate tolDCs ex vivo and common features shared by the tolDCs are shown in Fig. 1.
Pharmacological agents
Pharmacological agents known to induce tolDCs include vitamin D3, corticosteroid, rapamycin, cyclosporine, tacrolimus, aspirin, atorvastatin, retinoic acid, mycophenolic acid, and minocycline.1011161718192021 Of these agents, vitamin D3, dexamethasone, and rapamycin have been extensively studied in experimental animals and in humans with the aim of developing clinical approaches for the prevention of transplantation rejection and treatment of autoimmune and chronic inflammatory conditions.
The biologically active form of vitamin D, 1,25-dihydroxyvitamin D3 [1,25(OH)2D3], is able to promote the generation of tolDCs.2223 DCs generated using vitamin D express lower levels of MHC class II and co-stimulatory molecules, and produce higher amount of IL-10 and lower amounts of IL-12 and IL-6, compared to untreated normal DCs.2223 Moreover, these DCs are poor activators of antigen-primed T cells, but stimulate the generation of Tregs.24 The tolDC-inducing activity of vitamin D has also been demonstrated in diabetes-prone NOD mice and normal mice.25
Corticosteroids, dexamethasone and prednisolone, have long been known to exert anti-inflammatory and immunosuppressive activities. Numerous studies have shown that corticosteroids exert their immunosuppressive activity at least in part via induction of tolDCs. DCs generated in the presence of dexamethasone express low levels of co-stimulatory molecules and MHC class II molecules, produce elevated levels of IL-10 and lower levels of IL-12, and induce the generation of Tregs.131626 Dexamethasone also induces the generation of tolerogenic macrophages.26 Moreover, DCs generated with dexamethasone retain their tolerogenicity for several days, up to a week, even after dexamethasone is removed.1626
Rapamycin has long been known to suppress T cell activation via inhibition of the serine/threonine protein kinase, mammalian target of rapamycin. Rapamycin also induces the generation of tolDCs. DCs generated with rapamycin are poor stimulators of antigen-primed T cells, resistant to maturation induced by anti-CD40 or lipopolysaccharide (LPS) stimulation, and enhance the generation of Foxp3+ Tregs.27282930 Treatment of murine heart transplantation recipients with rapamycin-generated DCs increases the survival of the transplanted organ, in correlation with increased production of Foxp3+ Tregs in the recipient mice.27
One of the drawbacks of generating tolDCs using the above listed pharmacologic agents is the cytotoxic effects of these drugs. For instance, rapamycin (10 ng/mL) is effective in generating tolDCs from bone marrow cells when used together with GM-CSF and IL-4. However, the number of CD11c+ cells obtained from rapamycin-conditioned cultures is significantly (more than 40%) lower than that from rapamycin-unconditioned cultures.27 Dexamethasone has also been shown to markedly reduce DC recovery.1626 In this regard, minocycline is unique in that it increases the generation of tolDCs from bone marrow cells.21 Minocycline also exerts growth-promoting effects on DCs conditioned with relatively toxic doses of rapamycin, vitamin D3, or IL-10.31 Furthermore, the tolerogenicity of tolDCs generated in the presence of minocycline and dexamethasone is superior or at least equal to that of tolDCs generated with either one of these agents.31
Combinations of pharmacological agents are also used to generate tolDCs with potent tolerogenic properties. For instance, potently tolerogenic and highly stable tolDCs are generated from monocytes of rheumatoid arthritis (RA) patients by the addition of dexamethasone, vitamin D3, and monophosphoryl lipid A together with GM-CSF and IL-4.32
Immunosuppressive cytokines
Immunosuppressive cytokines such as IL-10 and TGF-β have been shown to induce regulatory DCs.333435 Other cytokines known to induce tolDCs include TNF-α,36 interferon (IFN)-γ,37 hepatocyte growth factor,38 and IL-21.39 TolDCs generated with IL-10 have been extensively studied in experimental animals and in humans.
DCs generated with IL-10 display reduced levels of MHC class II molecules and co-stimulatory molecules, and induce the generation of Tregs.3334354041 DCs generated with IL-10 secrete high levels of IL-10 in the absence of IL-12.34 A comparative study demonstrated that the tolerogenic properties of IL-10-generated DCs are superior to those of vitamin D3-, dexamethasone-, or rapamycin-generated DCs.40 In addition, IL-10 in combination with TGF-β induce the generation of tolDCs with potent tolerogenic properties.
The fact that IFN-γ, a prototype of the Th1-type cytokine produced mainly by natural killer (NK) and T cells, induces the generation of tolDCs is somewhat surprising. At a low dose, IFN-γ promotes the maturation of DCs with full activating potential, however, a high dose of IFN-γ induces DC acquisition of regulatory features.42 The importance of the timing and intensity of IFN-γ exposure for the function of monocyte-derived DCs (mo-DCs) was also noted in a separate study.43 A dose-dependent and bivalent effect of IFN-γ on DC function would constitue a novel mechanism for homeostatic regulation of immune responses at local sites.
Genetic modifications
Genetic engineering of DCs to express immunosuppressive molecules is a method for generating tolDCs. DCs engineered to express IL-10 using a retroviral vector exhibit significantly reduced capacity to induce allogeneic T cell proliferation and cytotoxic T lymphocyte (CTL) generation.44 Over expression of TGF-β also promotes the tolerogenic potential of the DCs.45 DCs transduced with cDNA encoding CTLA-4-Ig demonstrate markedly reduced expression of co-stimulatory molecule CD86, but not MHC class II molecules, and induce antigen-specific hyporesponsiveness.46 DCs engineered to express indoleamine 2,3-dioxygenase (IDO) or Fas ligand (FasL) also exhibit tolerogenic properties.4748
Modulation of microRNA expression in DCs is another approach for generating tolDCs. Inhibition of miRNA let-7i in DCs results in low surface expression of co-stimulatory molecules, impaired T cell stimulatory capacity, and promotion of Treg induction.49 DCs transfected with miR-23b show decreased antigen uptake, increased IL-10 production, decreased IL-12 production, and an enhanced capacity to promote Treg differentiation.50
CARACTERISTICS AND MECHANISMS OF tolDC TOLEROGENICITY
The mechanisms by which tolDCs exert their activity are varied and incompletely understood. Moreover, phenotypic and functional differences among tolDCs arise intrinsically because of differences in the methods used to generate them. Nevertheless, there are common features shared by tolDCs, they exert an immature phenotype, and are resistant to maturation stimuli. The major mechanisms underlying the tolerance-inducing activity of tolDCs are reduction of co-stimulatory molecules, expression of various co-inhibitory molecules, production of immunosuppressive cytokines and mediators, and induction of Tregs.
Reduced expression of co-stimulatory molecules
The interaction of co-stimulatory molecules, such as CD80 and CD86, on DCs with CD28 on T cells triggers a T cell-activating signal. It is generally accepted that T cells become anergic and lose their ability to proliferate during subsequent stimulation when they are stimulated with signal-1, the recognition of MHC-complexed antigenic peptide via T cell receptor, in the absence of signals delivered from CD80 and CD86.2 Reduction of the expression levels of co-stimulatory molecules is one of the hallmarks of tolDCs, regardless of the methods used to generate them. TolDCs lacking co-stimulatory molecules induce T cell anergy.345
Increased expression of co-inhibitory molecules
TolDCs express increased levels of various co-inhibitory molecules such as PD-L1 and ICOSL.121351 T cells become functionally inactive following their interaction with co-inhibitory molecules. A number of tolDCs also express inhibitory Ig-like transcripts (ILTs) on their surface, which interact with MHC-I molecules, especially human leukocyte antigen (HLA)-G, and deliver negative signals to T cells. ILT3 and ILT4 are upregulated by exposing immature DCs to known immunosuppressive factors such as IL-10 and vitamin D3.5253
Production of immunosuppressive cytokines and mediators
Production of immunosuppressive cytokines, such as IL-10 and/or TGF-β, is one of the most common features of tolDCs.11 These cytokines inhibit the production of inflammatory cytokines, such as IL-12, TNF-α, and IFN-γ, and impairs the activation of T cells and NK cells.54 In addition, these cytokines induce Treg generation. IL-10, in particular, is crucial for the induction of IL-10-secreting T regulatory type 1 cells (Tr-1) cells.5556 Other immunosuppressive mediators known to be produced by tolDCs include IDO, hemoxygenase-1, and FasL. IDO has been known to suppress T and NK cells, and also induces Treg generation.575859 FasL-expressing tolDCs induce T cell apoptosis via the Fas/FasL interaction pathways.
Induction of Tregs
The ability of tolDCs to direct T cell polarization toward various types of Tregs is pivotal to their tolerogenic function. TolDCs induce several subtypes of regulatory lymphocytes such as CD4+CD25+Foxp3+ Tregs, CD25+Foxp3+/− Tr-1 cells, CD8+ Tregs, and regulatory B cells.27606162 CD4+CD25+Foxp3+ Tregs have been extensively investigated in various inflammatory diseases.636465 IL-10 and TGF-β are the major cytokines produced by tolDCs and induce Treg generation. IL-10-induced tolDCs acquire the ability to secrete IL-10, which exerts powerful anti-inflammatory effects and contributes to Treg differentiation and proliferation.66 TGF-β is unique among cytokines in that it induces Foxp3 expression and promotes Treg differentiation even in the absence of DCs.67 Foxp3+ Tregs, in turn, augments the generation and tolerogenic properties of tolDCs by suppressing DC maturation.6869
CLINICAL TRIALS WITH tolDCs
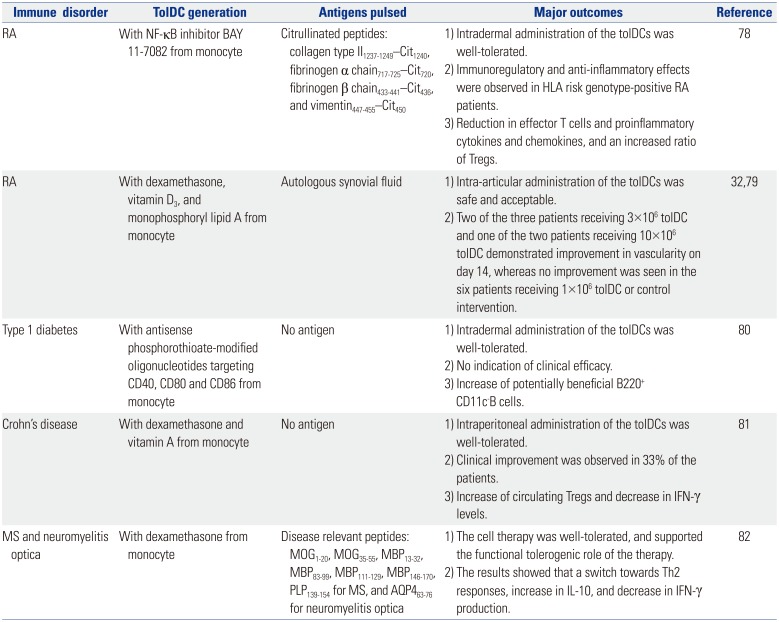
The therapeutic efficacy of ex vivo-generated tolDCs has been demonstrated in animal models of autoimmune diseases such as RA,707172 diabetes,7374 and experimental allergic encephalomyelitis,75 as well as in animal models of graft rejection.7677 Based on the successes in small animal models, several clinical trials have been completed or on-going in patients with autoimmune diseases such as RA, type 1 diabetes, multiple sclerosis (MS), and Crohn's disease. Five complete clinical trials with reported outcomes are summarized in Table 1.
The first trial for RA treatment was performed with tolDCs generated from monocytes by adding the NF-κB inhibitor, BAY 11-7082.78 The tolDCs were exposed to four citrullinated peptide antigens, collagen type II1237–1249—Cit1240, fibrinogen α chain717–725—Cit720, fibrinogen β chain433–441—Cit436, and vimentin447–455—Cit450, and then administered once via intradermal injection. The results showed that a single intradermal injection of tolDCs was safe, and effective in HLA risk genotype-positive RA patients. Another clinical trial for RA treatment was performed with tolDCs generated from monocytes by adding dexamethasone, vitamin D3, and monophosphoryl lipid A.3279 Antigens in autologous synovial fluid were loaded with these tolDCs and then administered into an inflamed knee joint via intra-articular injection. The treatment was deemed safe and acceptable with promising outcomes. Two of the three patients receiving 3×106 tolDCs and one of the two patients receiving 10×106 tolDCs demonstrated improvement in vascularity on day 14, whereas no improvement was observed in the six patients receiving 1×106 tolDCs or the control intervention.
Two clinical trials were performed with tolDCs not loaded with particular antigens. The phase I study of autologous tolDCs in type 1 diabetes patients was performed with tolDCs generated from monocytes with antisense oligonucleotides targeting the primary transcripts of the CD40, CD80 and CD86 co-stimulatory molecules.80 The treatment appeared safe and well tolerated. However, there was no indication of efficacy, although an increase in potentially beneficial B220+ CD11c− B cells was observed. Another clinical trial with antigen-unloaded tolDCs was performed in patients with Crohn's disease.81 In this study, tolDCs were generated from monocytes using dexamethasone and vitamin A. The treatment appeared safe and well tolerated, and resulted in clinical improvement in 33% of the patients.
Recently, a phase 1b clinical trial with antigen-loaded tolDCs was completed in patients with MS and neuromyelitis optica.82 In this study, tolDCs were generated from monocytes with dexamethasone, and loaded with disease relevant peptides, i.e., MOG1–20, MOG35–55, MBP13–32, MBP83–99, MBP111–129, MBP146–170, PLP139–154 for MS and AQP463–76 for neuromyelitis optica. The treatment was well-tolerated, and supported the functional tolerogenic efficacy of the therapy as demonstrated by a switch towards Th2 responses, an increase in IL-10 production, and a decrease in IFN-γ production.
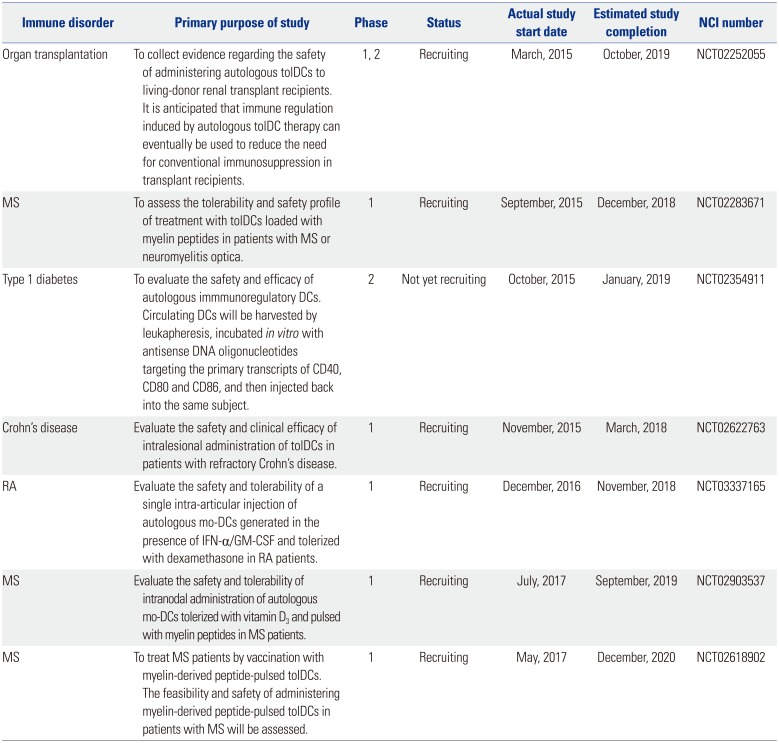
The primary purposes of the clinical trials currently registered with ClinicalTrials.gov are summarized in Table 2. The current on-going clinical trials with tolDCs mostly involve autoimmune diseases such as Crohn's disease, RA, MS, and type 1 diabetes mellitus. One clinical trial aiming to reduce the need for conventional immunosuppression in transplant recipients is also underway. As shown in Table 2, different approaches are being used to generate tolDCs for clinical use, raising the need to establish a more standardized ex vivo generation method(s). In fact, there are numerous questions that have to be addressed in order to achieve generalized and successful application of tolDCs in clinical settings including optimization of dose, route, and frequency of administration. Nevertheless, the list in Table 2 show that tolDC-based treatment of autoimmune diseases is now a reality, and could constitute an innovative cellular therapy in the future.
CONCLUSION AND PROSPECTS
TolDCs can be generated ex vivo from peripheral blood monocytes or bone marrow cells by culturing them in the presence of GM-CSF, IL-4, and an agent(s) known to confer tolerogenic properties. The agents used extensively to generate tolDCs include vitamin D, dexamethasone, rapamycin, and IL-10, and new agents, such as minocycline, are being continuously explored. Although the mechanisms by which tolDCs exert their activity are diverse and incompletely understood, there are common features shared by tolDCs. In general, tolDCs exert an immature phenotype, and are resistant to maturation stimuli. TolDCs are characterized by reduced expression of co-stimulatory molecules, increased expression of co-inhibitory molecules, production of immunosuppressive cytokines and mediators, and/or induction of Tregs. Based on the successes in small animal models, several clinical trials have been completed or are on-going in patients with autoimmune diseases such as RA, type 1 diabetes, MS, and Crohn's disease. The results thus far are highly encouraging both in terms of safety and clinical efficacy in all the clinical studies completed to date, tolDC administration is tolerated and appears safe. More importantly, the completed clinical trials indicate significant promise for tolDC-based immunotherapy. However, numerous questions remain to be addressed prior to generalized and successful application of tolDCs in clinical settings. One of the major challenges facing tolDC-based immunotherapy is protocol optimization in order to obtain a maximum number of tolDCs with stable tolerogenic properties. In addition, the dose, route, and frequency of administration of each type of tolDC also require optimization. However, as demonstrated by numerous on-going clinical studies, tolDC-based treatment of autoimmune diseases is now a reality, and could provide innovative cellular therapy in the future.
 XML Download
XML Download